
Biochemia bólu:„Wszystkie zespoły bólowe mają profil zapalny. Profil stanu zapalnego może się różnić w zależności od osoby, a także może różnić się u jednej osoby w różnym czasie. Leczenie zespołów bólowych polega na zrozumieniu tego profilu zapalnego. Zespoły bólowe są leczone medycznie, chirurgicznie lub jedno i drugie. Celem jest zahamowanie/zahamowanie produkcji mediatorów stanu zapalnego. A pomyślny wynik to taki, który skutkuje mniejszym stanem zapalnym i oczywiście mniejszym bólem.
Spis treści
Biochemia bólu
Cele:
- Kim są główni gracze?
- Jakie są mechanizmy biochemiczne?
- Jakie są konsekwencje?
Przegląd stanu zapalnego:
Kluczowi gracze
 Dlaczego boli mnie ramię? Przegląd neuroanatomicznej i biochemicznej podstawy bólu barku
Dlaczego boli mnie ramię? Przegląd neuroanatomicznej i biochemicznej podstawy bólu barku
ABSTRACT
Jeśli pacjent zapyta „dlaczego boli mnie ramię?”, rozmowa szybko przerodzi się w teorię naukową, a czasem bezpodstawne domysły. Często klinicysta uświadamia sobie ograniczenia naukowej podstawy ich wyjaśnienia, wykazując niekompletność naszego rozumienia natury bólu barku. W niniejszym przeglądzie przyjęto systematyczne podejście, aby pomóc odpowiedzieć na podstawowe pytania dotyczące bólu barku, z myślą o zapewnieniu wglądu w przyszłe badania i nowe metody leczenia bólu barku. Zbadamy role (1) receptorów obwodowych, (2) przetwarzania bólu obwodowego lub „nocycepcji”, (3) rdzenia kręgowego, (4) mózgu, (5) lokalizacji receptorów w barku i (6 ) anatomia nerwowa barku. Zastanawiamy się również, w jaki sposób czynniki te mogą przyczynić się do zmienności obrazu klinicznego, diagnozy i leczenia bólu barku. W ten sposób zamierzamy przedstawić przegląd elementów składowych systemu wykrywania bólu obwodowego i ośrodkowych mechanizmów przetwarzania bólu w bólu barku, które współdziałają w celu wywołania bólu klinicznego.
WPROWADZENIE: BARDZO KRÓTKA HISTORIA NAUKI BÓLU NIEZBĘDNEJ DLA KLINIKÓW
Ogólnie rzecz biorąc, natura bólu była przedmiotem wielu kontrowersji w ciągu ostatniego stulecia. W XVII-wiecznej teorii Kartezjusza17 sugerowano, że intensywność bólu jest bezpośrednio związana z rozmiarem uszkodzenia tkanki towarzyszącej i że ból jest przetwarzany jedną odrębną ścieżką. Wiele wcześniejszych teorii opierało się na tej tak zwanej „dualistycznej” filozofii kartezjańskiej, widząc ból jako konsekwencję stymulacji „specyficznego” obwodowego receptora bólu w mózgu. W XX wieku wywiązała się naukowa bitwa pomiędzy dwiema przeciwstawnymi teoriami, mianowicie teorią specyficzności i teorią wzorców. Kartezjańska „teoria specyficzności” postrzegała ból jako specyficzną, oddzielną modalność bodźców sensorycznych z własnym aparatem, podczas gdy „teoria wzorców” uważała, że ból jest wynikiem intensywnej stymulacji niespecyficznych receptorów.1 W 20 Wall i Melzack 2 teoria bramkowa bólu dostarczyła dowodów na istnienie modelu, w którym percepcja bólu była modulowana zarówno przez czuciowe sprzężenie zwrotne, jak i centralny układ nerwowy. Kolejnym ogromnym postępem w teorii bólu, mniej więcej w tym samym czasie, było odkrycie specyficznego sposobu działania opioidów.1965 Następnie ostatnie postępy w neuroobrazowaniu i medycynie molekularnej znacznie poszerzyły naszą ogólną wiedzę na temat bólu.
Więc jak to się ma do bólu barku?�Ból barku jest częstym problemem klinicznym, a dogłębne zrozumienie sposobu, w jaki ból jest przetwarzany przez organizm, jest niezbędne do najlepszego diagnozowania i leczenia bólu pacjenta. Postępy w naszej wiedzy na temat przetwarzania bólu obiecują wyjaśnić rozbieżność między patologią a odczuwaniem bólu, mogą również pomóc nam wyjaśnić, dlaczego niektórzy pacjenci nie reagują na określone terapie.
PODSTAWOWE BUDOWNICTWO BÓLU
Obwodowe receptory czuciowe: mechanoreceptor i „nocyceptor”
W ludzkim układzie mięśniowo-szkieletowym występuje wiele rodzajów obwodowych receptorów czuciowych. 5 Mogą być klasyfikowane na podstawie ich funkcji (jako mechanoreceptory, termoreceptory lub nocyceptory) lub morfologii (wolne zakończenia nerwowe lub różne typy receptorów zamkniętych w kapsułce).5 Różne typy receptorów można następnie dalej klasyfikować w oparciu o obecność niektórych markerów chemicznych. Istnieją znaczne nakładania się różnych klas funkcjonalnych receptora, na przykład:
Przetwarzanie bólu obwodowego: „Nocycepcja”
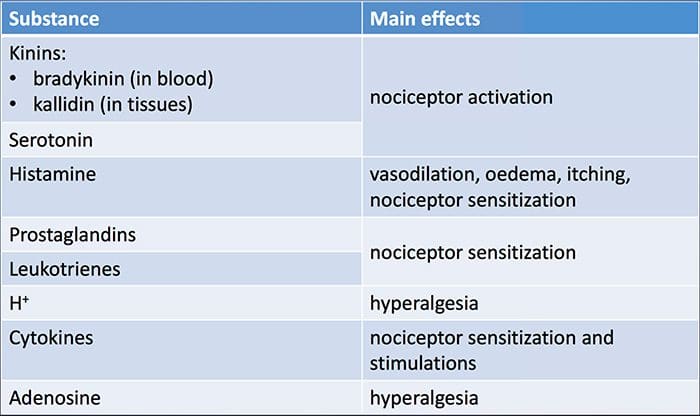
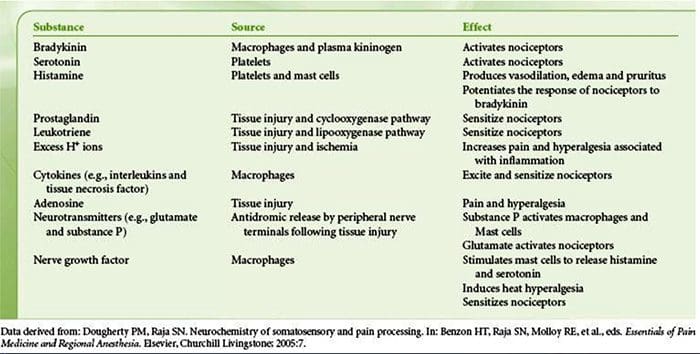
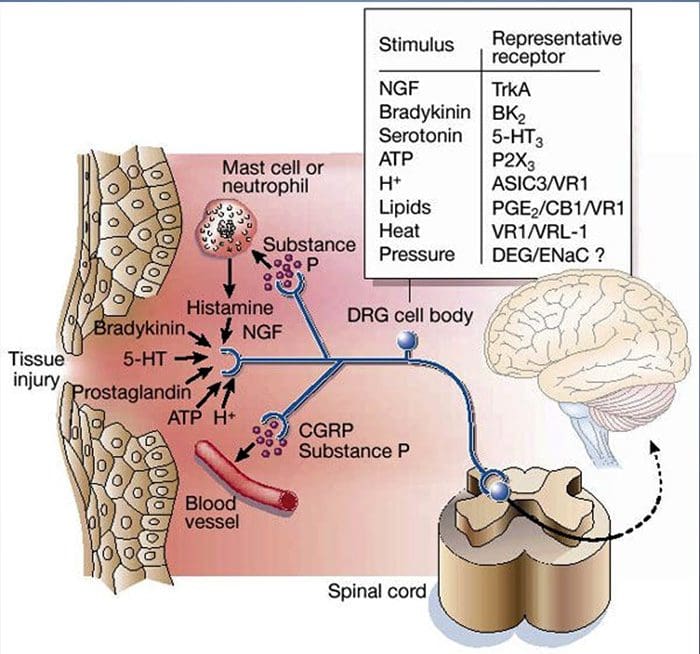
Uszkodzenie tkanki obejmuje różne mediatory zapalne uwalniane przez uszkodzone komórki, w tym bradykininę, histaminę, 5-hydroksytryptaminę, ATP, tlenek azotu i niektóre jony (K+ i H+). Aktywacja szlaku kwasu arachidonowego prowadzi do produkcji prostaglandyn, tromboksanów i leukotrienów. Cytokiny, w tym interleukiny i czynnik martwicy nowotworu a, oraz neurotrofiny, takie jak czynnik wzrostu nerwów (NGF), są również uwalniane i są ściśle zaangażowane w wywoływanie stanu zapalnego.15 Inne substancje, takie jak aminokwasy pobudzające (glutaminian) i opioidy ( endotelina-1) była również zaangażowana w ostrą odpowiedź zapalną.16 17 Niektóre z tych czynników mogą bezpośrednio aktywować nocyceptory, podczas gdy inne powodują rekrutację innych komórek, które następnie uwalniają kolejne czynniki ułatwiające.18 Ten lokalny proces skutkuje zwiększoną reaktywnością. neuronów nocyceptywnych do ich normalnego sygnału wejściowego i/lub rekrutacja odpowiedzi na sygnały normalnie podprogowe jest określana jako „uczulenie obwodowe”. Rysunek 1 podsumowuje niektóre kluczowe mechanizmy.
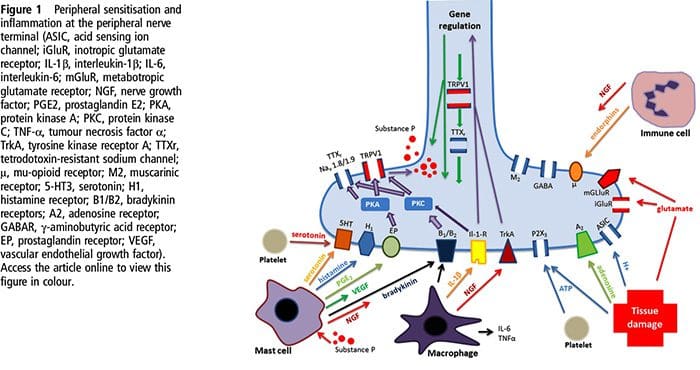 NGF i receptor członu 1 kanału kationowego podrodziny V podrodziny receptora przejściowego potencjalnego receptora (TRPV1) mają symbiotyczny związek, jeśli chodzi o zapalenie i uczulenie na receptory nocyceptorowe. Cytokiny wytwarzane w tkance w stanie zapalnym powodują wzrost produkcji NGF.19 NGF stymuluje uwalnianie histaminy i serotoniny (5-HT3) przez komórki tuczne, a także uwrażliwia nocyceptory, prawdopodobnie zmieniając właściwości A? włókna tak, że większa część staje się nocyceptywna. Receptor TRPV1 jest obecny w subpopulacji pierwotnych włókien doprowadzających i jest aktywowany przez kapsaicynę, ciepło i protony. Receptor TRPV1 jest syntetyzowany w ciele komórki włókna aferentnego i jest transportowany zarówno do terminali obwodowych, jak i centralnych, gdzie przyczynia się do wrażliwości neuronów aferentnych nocyceptywnych. Zapalenie powoduje obwodowe wytwarzanie NGF, który następnie wiąże się z receptorem kinazy tyrozynowej typu 1 na końcach nocyceptorów, NGF jest następnie transportowany do ciała komórki, gdzie prowadzi do regulacji w górę transkrypcji TRPV1, a w konsekwencji do zwiększonej wrażliwości na receptory nocyceptorowe.19 20 NGF i inne mediatory zapalne również uwrażliwiają TRPV1 poprzez różnorodną gamę drugorzędowych szlaków przekaźnikowych. Uważa się, że wiele innych receptorów, w tym receptory cholinergiczne, receptory kwasu y-aminomasłowego (GABA) i receptory somatostatyny, są również zaangażowane w wrażliwość obwodowych receptorów nocyceptorowych.
NGF i receptor członu 1 kanału kationowego podrodziny V podrodziny receptora przejściowego potencjalnego receptora (TRPV1) mają symbiotyczny związek, jeśli chodzi o zapalenie i uczulenie na receptory nocyceptorowe. Cytokiny wytwarzane w tkance w stanie zapalnym powodują wzrost produkcji NGF.19 NGF stymuluje uwalnianie histaminy i serotoniny (5-HT3) przez komórki tuczne, a także uwrażliwia nocyceptory, prawdopodobnie zmieniając właściwości A? włókna tak, że większa część staje się nocyceptywna. Receptor TRPV1 jest obecny w subpopulacji pierwotnych włókien doprowadzających i jest aktywowany przez kapsaicynę, ciepło i protony. Receptor TRPV1 jest syntetyzowany w ciele komórki włókna aferentnego i jest transportowany zarówno do terminali obwodowych, jak i centralnych, gdzie przyczynia się do wrażliwości neuronów aferentnych nocyceptywnych. Zapalenie powoduje obwodowe wytwarzanie NGF, który następnie wiąże się z receptorem kinazy tyrozynowej typu 1 na końcach nocyceptorów, NGF jest następnie transportowany do ciała komórki, gdzie prowadzi do regulacji w górę transkrypcji TRPV1, a w konsekwencji do zwiększonej wrażliwości na receptory nocyceptorowe.19 20 NGF i inne mediatory zapalne również uwrażliwiają TRPV1 poprzez różnorodną gamę drugorzędowych szlaków przekaźnikowych. Uważa się, że wiele innych receptorów, w tym receptory cholinergiczne, receptory kwasu y-aminomasłowego (GABA) i receptory somatostatyny, są również zaangażowane w wrażliwość obwodowych receptorów nocyceptorowych.
Duża liczba mediatorów zapalnych jest szczególnie związana z bólem barku i chorobą stożka rotatorów.21 Podczas gdy niektóre mediatory chemiczne bezpośrednio aktywują nocyceptory, większość z nich prowadzi do zmian w samym neuronie czuciowym, a nie bezpośrednio go aktywuje. Zmiany te mogą być zależne od wczesnej potranslacyjnej lub opóźnionej transkrypcji. Przykładami tych pierwszych są zmiany w receptorze TRPV25 lub w kanałach jonowych bramkowanych napięciem, wynikające z fosforylacji białek związanych z błoną. Przykłady tych ostatnich obejmują indukowany przez NGF wzrost wytwarzania kanału TRV1 i indukowaną wapniem aktywację wewnątrzkomórkowych czynników transkrypcyjnych.
Molekularne mechanizmy nocycepcji
Uczucie bólu ostrzega nas o prawdziwym lub zbliżającym się urazie i wyzwala odpowiednie reakcje ochronne. Niestety ból często przeżywa swoją przydatność jako system ostrzegawczy i zamiast tego staje się chroniczny i wyniszczający. To przejście do fazy przewlekłej obejmuje zmiany w rdzeniu kręgowym i mózgu, ale istnieje również niezwykła modulacja, w której inicjowane są komunikaty bólowe – na poziomie pierwotnego neuronu czuciowego. Wysiłki zmierzające do ustalenia, w jaki sposób te neurony wykrywają bodźce wywołujące ból o charakterze termicznym, mechanicznym lub chemicznym, ujawniły nowe mechanizmy sygnalizacji i przybliżyły nas do zrozumienia zdarzeń molekularnych, które ułatwiają przejście od bólu ostrego do uporczywego.
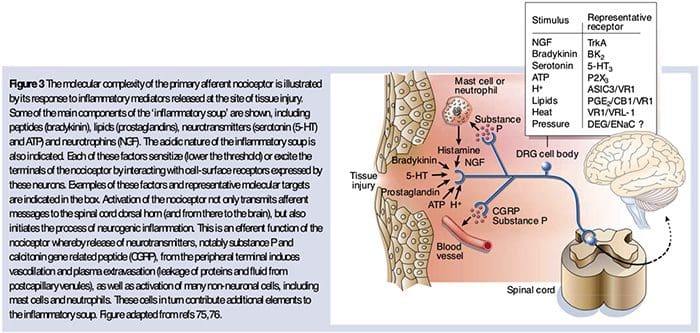 Neurochemia nocyceptorów
Neurochemia nocyceptorów
Glutaminian jest głównym neuroprzekaźnikiem pobudzającym we wszystkich nocyceptorach. Badania histochemiczne dorosłych DRG ujawniły jednak dwie szerokie klasy niezmielinizowanego włókna C.
Przetworniki chemiczne, aby pogorszyć ból
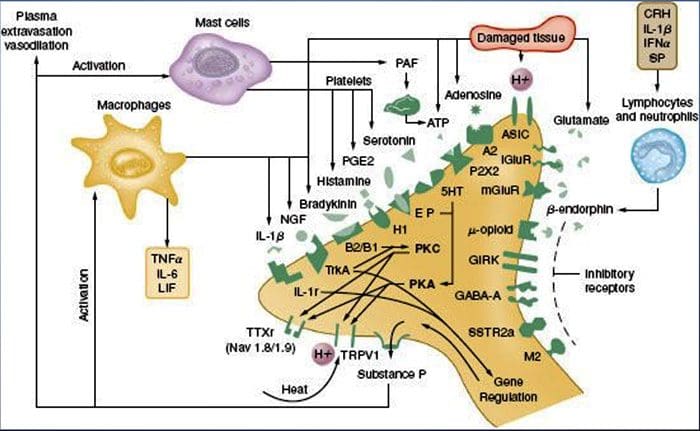
Jak opisano powyżej, uraz zwiększa nasze odczuwanie bólu poprzez zwiększenie wrażliwości nocyceptorów na bodźce zarówno termiczne, jak i mechaniczne. Zjawisko to wynika po części z wytwarzania i uwalniania mediatorów chemicznych z pierwotnego terminala czuciowego oraz z komórek nienerwowych (np. fibroblastów, komórek tucznych, neutrofili i płytek krwi) w środowisku36 (ryc. 3). Niektóre składniki zupy zapalnej (na przykład protony, ATP, serotonina lub lipidy) mogą bezpośrednio zmieniać pobudliwość neuronów poprzez interakcję z kanałami jonowymi na powierzchni nocyceptora, podczas gdy inne (na przykład bradykinina i NGF) wiążą się z receptorami metabotropowymi i pośredniczą w ich skutkach poprzez kaskady sygnalizacyjne drugiego posłańca11. Poczyniono znaczne postępy w zrozumieniu podstaw biochemicznych takich mechanizmów modulujących.
Protony zewnątrzkomórkowe i kwasica tkankowa
Miejscowa kwasica tkankowa jest charakterystyczną fizjologiczną reakcją na uraz, a stopień związanego z nią bólu lub dyskomfortu jest dobrze skorelowany ze stopniem zakwaszenia37. Podawanie kwasu (pH 5) na skórę powoduje długotrwałe wyładowania w co najmniej jednej trzeciej nocyceptorów polimodalnych, które unerwiają pole recepcyjne 20.
 Komórkowe i molekularne mechanizmy bólu
Komórkowe i molekularne mechanizmy bólu
Abstrakcyjny
Układ nerwowy wykrywa i interpretuje szeroką gamę bodźców termicznych i mechanicznych, a także środowiskowe i endogenne chemiczne czynniki drażniące. Gdy są intensywne, bodźce te generują ostry ból, aw sytuacji trwałego urazu, zarówno obwodowe, jak i ośrodkowe elementy układu nerwowego szlaku przekazywania bólu wykazują ogromną plastyczność, wzmacniając sygnały bólowe i wywołując nadwrażliwość. Kiedy plastyczność ułatwia odruchy ochronne, może to być korzystne, ale gdy zmiany utrzymują się, może wystąpić przewlekły ból. Badania genetyczne, elektrofizjologiczne i farmakologiczne wyjaśniają mechanizmy molekularne, które leżą u podstaw wykrywania, kodowania i modulowania szkodliwych bodźców wywołujących ból.
Wstęp: ostry kontra uporczywy ból
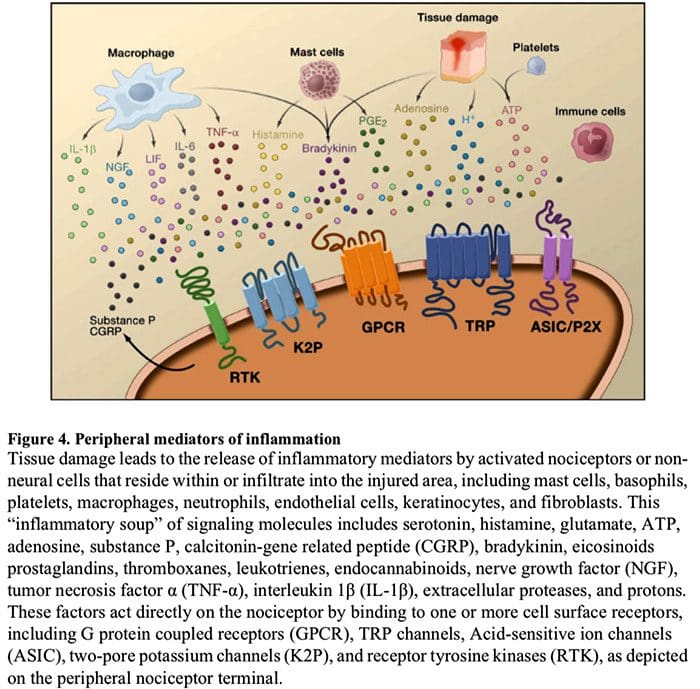
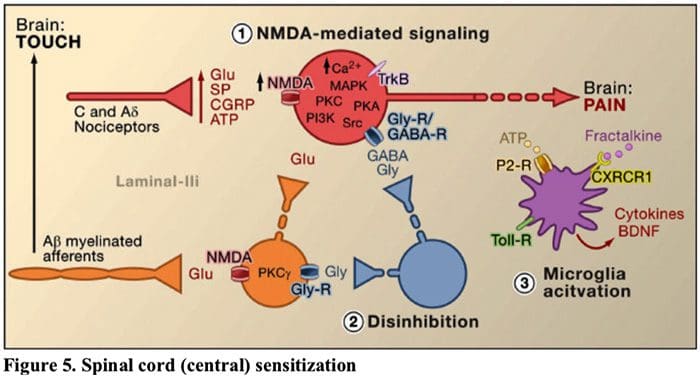 Rycina 5. Uczulenie rdzenia kręgowego (ośrodkowe)
Rycina 5. Uczulenie rdzenia kręgowego (ośrodkowe)
- Uczulenie za pośrednictwem receptora glutaminianu/NMDA.�Po intensywnej stymulacji lub uporczywym urazie, aktywowane C i A? nocyceptory uwalniają różne neuroprzekaźniki, w tym dlutaminian, substancję P, peptyd związany z genem kalcytoniny (CGRP) i ATP, do neuronów wyjściowych w blaszce I powierzchownego rogu grzbietowego (czerwony). W konsekwencji normalnie ciche receptory glutaminianu NMDA zlokalizowane w neuronie postsynaptycznym mogą teraz sygnalizować, zwiększać poziom wapnia wewnątrzkomórkowego i aktywować wiele ścieżek sygnałowych zależnych od wapnia i wtórnych przekaźników, w tym kinazę białkową aktywowaną mitogenem (MAPK), kinazę białkową C (PKC) , kinaza białkowa A (PKA) i Src. Ta kaskada zdarzeń zwiększy pobudliwość neuronu wyjściowego i ułatwi przekazywanie wiadomości bólowych do mózgu.
- Odhamowanie.„W normalnych warunkach interneurony hamujące (niebieskie) w sposób ciągły uwalniają GABA i/lub glicynę (Gly), aby zmniejszyć pobudliwość neuronów wyjściowych blaszki I i modulować przenoszenie bólu (ton hamujący). Jednak w sytuacji urazu hamowanie to może zostać utracone, co prowadzi do przeczulicy bólowej. Dodatkowo, odhamowanie może umożliwić nienocyceptywną mielinizację A? pierwotne aferenty angażujące obwody transmisji bólu tak, że normalnie nieszkodliwe bodźce są teraz odbierane jako bolesne. Dzieje się to po części poprzez odhamowanie pobudzającego PKC? wyrażanie interneuronów w wewnętrznej blaszce II.
- Aktywacja mikrogleju.„Uszkodzenie nerwów obwodowych sprzyja uwalnianiu ATP i chemokiny fraktalkiny, które będą stymulować komórki mikrogleju. W szczególności aktywacja receptorów purynergicznych, CX3CR1 i Toll-podobnych na mikrogleju (fioletowy) powoduje uwolnienie neurotroficznego czynnika pochodzenia mózgowego (BDNF), który poprzez aktywację receptorów TrkB wyrażanych przez neurony wyprowadzające blaszkę I, sprzyja zwiększonej pobudliwości i nasilony ból w odpowiedzi zarówno na szkodliwą, jak i nieszkodliwą stymulację (tj. przeczulicę bólową i allodynię). Aktywowany mikroglej uwalnia również wiele cytokin, takich jak czynnik martwicy nowotworu? (TNF?), interleukina-1? i 6 (IL-la, IL-1) oraz inne czynniki, które przyczyniają się do uczulenia ośrodkowego.
Chemiczne środowisko zapalenia
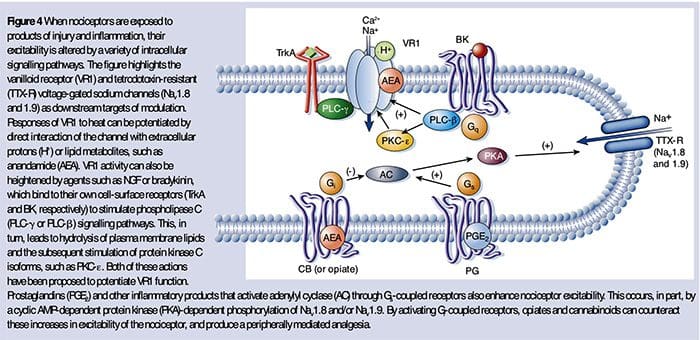
Uczulenie obwodowe częściej wynika ze zmian związanych ze stanem zapalnym w środowisku chemicznym włókna nerwowego (McMahon i wsp., 2008). Tak więc uszkodzeniu tkanki często towarzyszy akumulacja endogennych czynników uwalnianych z aktywowanych nocyceptorów lub komórek nieneuronalnych, które znajdują się w uszkodzonym obszarze lub przenikają do niego (w tym komórki tuczne, bazofile, płytki krwi, makrofagi, neutrofile, komórki śródbłonka, keratynocyty i fibroblasty). Zbiorowo. czynniki te, określane mianem „zupy zapalnej”, reprezentują szeroki wachlarz cząsteczek sygnałowych, w tym neuroprzekaźniki, peptydy (substancja P, CGRP, bradykinina), eikozynoidy i pokrewne lipidy (prostaglandyny, tromboksany, leukotrieny, endokannabinoidy), neurotrofiny, cytokiny oraz chemokiny, a także proteazy zewnątrzkomórkowe i protony. Co godne uwagi, nocyceptory wyrażają jeden lub więcej receptorów powierzchniowych komórek zdolnych do rozpoznawania i odpowiadania na każdy z tych czynników prozapalnych lub proalgetycznych (Figura 4). Takie interakcje zwiększają pobudliwość włókna nerwowego, zwiększając w ten sposób jego wrażliwość na temperaturę lub dotyk.
Niewątpliwie najczęstsze podejście do zmniejszania bólu zapalnego polega na hamowaniu syntezy lub akumulacji składników zapalnej zupy. Najlepszym przykładem są niesteroidowe leki przeciwzapalne, takie jak aspiryna lub ibuprofen, które zmniejszają ból zapalny i przeczulicę bólową poprzez hamowanie cyklooksygenaz (Cox-1 i Cox-2) zaangażowanych w syntezę prostaglandyn. Drugie podejście polega na zablokowaniu działania czynników zapalnych w nocyceptorze. W tym miejscu przedstawiamy przykłady, które zapewniają nowy wgląd w komórkowe mechanizmy uczulenia obwodowego lub które stanowią podstawę nowych strategii terapeutycznych w leczeniu bólu zapalnego.
NGF jest prawdopodobnie najlepiej znany ze swojej roli jako czynnika neurotroficznego wymaganego do przeżycia i rozwoju neuronów czuciowych podczas embriogenezy, ale u dorosłych NGF jest również wytwarzany w warunkach uszkodzenia tkanki i stanowi ważny składnik zupy zapalnej (Ritner et al. al., 2009). Wśród wielu celów komórkowych, NGF działa bezpośrednio na nocyceptory peptydergicznych włókien C, które wyrażają kinazę tyrozynową receptora NGF o wysokim powinowactwie, TrkA, jak również receptor neurotrofiny o niskim powinowactwie, p75 (Chao, 2003; Snider i McMahon, 1998). NGF wytwarza głęboką nadwrażliwość na ciepło i bodźce mechaniczne dzięki dwóm czasowo odrębnym mechanizmom. Początkowo oddziaływanie NGF-TrkA aktywuje dalsze szlaki sygnałowe, w tym fosfolipazę C (PLC), kinazę białkową aktywowaną mitogenem (MAPK) i kinazę 3-fosfoinozytydu (PI3K). Powoduje to funkcjonalne wzmocnienie białek docelowych na obwodowym końcu nocyceptora, w szczególności TRPV1, prowadząc do szybkiej zmiany wrażliwości komórkowej i behawioralnej na ciepło (Chuang i wsp., 2001).
Niezależnie od ich mechanizmów pronocyceptywnych, zakłócanie sygnalizacji neurotrofin lub cytokin stało się główną strategią kontrolowania choroby zapalnej lub wynikającego z niej bólu. Główne podejście polega na blokowaniu NGF lub TNF-? działanie z neutralizującym przeciwciałem. W przypadku TNF-a, okazało się to niezwykle skuteczne w leczeniu licznych chorób autoimmunologicznych, w tym reumatoidalnego zapalenia stawów, prowadząc do dramatycznego zmniejszenia zarówno zniszczenia tkanki, jak i towarzyszącej hiperalgezji (Atzeni i wsp., 2005). Ponieważ główne działania NGF na receptory nocyceptorów dorosłych występują w warunkach zapalenia, zaletą tego podejścia jest zmniejszenie przeczulicy bólowej bez wpływu na normalne postrzeganie bólu. Rzeczywiście, przeciwciała anty-NGF są obecnie w badaniach klinicznych w leczeniu zespołów bólu zapalnego (Hefti i wsp., 2006).
Uczulenie za pośrednictwem receptora glutaminianu/NMDA
Ostry ból jest sygnalizowany uwalnianiem glutaminianu z centralnych końcówek nocyceptorów, generując pobudzające prądy postsynaptyczne (EPSC) w neuronach rogu grzbietowego drugiego rzędu. Dzieje się to głównie poprzez aktywację postsynaptycznych podtypów AMPA i kainowych jonotropowych receptorów glutaminianu. Sumowanie podprogowych EPSC w neuronie postsynaptycznym w końcu spowoduje odpalenie potencjału czynnościowego i przekazanie wiadomości o bólu do neuronów wyższego rzędu.
Inne badania wskazują, że same zmiany w neuronie projekcyjnym przyczyniają się do procesu odhamowania. Na przykład, uszkodzenie nerwów obwodowych głęboko obniża poziom transportera K+-Cl- KCC2, który jest niezbędny do utrzymania normalnych gradientów K+ i Cl- przez błonę plazmatyczną (Coull i wsp., 2003). Regulacja w dół KCC2, która jest wyrażana w neuronach projekcyjnych blaszki I, powoduje przesunięcie gradientu Cl-, tak że aktywacja receptorów GABA-A depolaryzuje, zamiast hiperpolaryzuje neurony projekcyjne blaszki I. To z kolei zwiększyłoby pobudliwość i zwiększyłoby przenoszenie bólu. Rzeczywiście, blokada farmakologiczna lub regulacja w dół KCC2 za pośrednictwem siRNA u szczura indukuje mechaniczną allodynię.

Udostępnij Ebook
Źródła:
Dlaczego boli mnie ramię? Przegląd neuroanatomicznych i biochemicznych podstaw bólu barku
Benjamina Johna Floyda Deana, Stephena Edwarda Gwilyma i Andrew Jonathana Carra
Komórkowe i molekularne mechanizmy bólu
Allan I. Basbaum1, Diana M. Bautista2, Gregory Scherrer1 i David Julius3
1Katedra Anatomii, Uniwersytet Kalifornijski, San Francisco 94158
2 Wydział Biologii Molekularnej i Komórkowej, Uniwersytet Kalifornijski, Berkeley CA 94720 3 Wydział Fizjologii Uniwersytetu Kalifornijskiego, San Francisco 94158
Molekularne mechanizmy nocycepcji
David Julius* i Allan I. Basbaum�
*Katedra Farmakologii Komórkowej i Molekularnej oraz Wydziały Anatomii i Fizjologii oraz WM Keck Foundation Center for Integrative Neuroscience, University of California San Francisco, San Francisco, California 94143, USA (e-mail: julius@socrates.ucsf.edu)
Profesjonalny zakres praktyki *
Informacje zawarte w niniejszym dokumencie na temat „Biochemia bólu„ nie ma na celu zastąpienia relacji jeden na jeden z wykwalifikowanym pracownikiem służby zdrowia lub licencjonowanym lekarzem i nie jest poradą medyczną. Zachęcamy do podejmowania decyzji dotyczących opieki zdrowotnej na podstawie badań i współpracy z wykwalifikowanym pracownikiem służby zdrowia.
Informacje o blogu i zakres dyskusji
Nasz zakres informacji ogranicza się do leków chiropraktycznych, mięśniowo-szkieletowych, fizykalnych, odnowy biologicznej, przyczyni się do etiologii zaburzenia trzewno-somatyczne w prezentacjach klinicznych, powiązanej dynamice klinicznej odruchu somato-trzewnego, kompleksach podwichnięć, wrażliwych kwestiach zdrowotnych i/lub artykułach, tematach i dyskusjach z zakresu medycyny funkcjonalnej.
Zapewniamy i prezentujemy współpraca kliniczna ze specjalistami z różnych dziedzin. Każdy specjalista podlega zakresowi swojej praktyki zawodowej i jurysdykcji licencyjnej. Stosujemy protokoły funkcjonalnego zdrowia i dobrego samopoczucia, aby leczyć i wspierać opiekę nad urazami lub zaburzeniami układu mięśniowo-szkieletowego.
Nasze filmy, posty, tematy, tematy i spostrzeżenia obejmują kwestie kliniczne, problemy i tematy, które bezpośrednio lub pośrednio odnoszą się do naszego zakresu praktyki klinicznej i wspierają ją.*
Nasze biuro podjęło zasadne próby dostarczenia wspierających cytatów i zidentyfikowało odpowiednie badanie lub badania wspierające nasze posty. Na żądanie udostępniamy kopie badań wspierających, które są dostępne dla organów regulacyjnych i opinii publicznej.
Rozumiemy, że zajmujemy się sprawami, które wymagają dodatkowego wyjaśnienia, w jaki sposób może to pomóc w konkretnym planie opieki lub protokole leczenia; w związku z tym, aby dokładniej omówić powyższy temat, prosimy o kontakt Dr Alex Jimenez, DC, lub skontaktować się z nami pod adresem 915-850-0900.
Jesteśmy tutaj, aby pomóc Tobie i Twojej rodzinie.
Błogosławieństwa
Dr Alex Jimenez OGŁOSZENIE, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionmedicine.com
Licencjonowany jako lekarz chiropraktyki (DC) w Texas & Nowy Meksyk*
Licencja Texas DC nr TX5807, Licencja DC w Nowym Meksyku nr NM-DC2182
Licencjonowana pielęgniarka dyplomowana (RN*) in Floryda
Licencja Florydy Licencja RN # RN9617241 (nr kontrolny 3558029)
Stan kompaktowy: Licencja wielostanowa: Uprawniony do wykonywania zawodu Stany 40*
Dr Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Moja cyfrowa wizytówka





