Back Clinic Wsparcie neurofizjologii klinicznej. El Paso, Teksas. Kręgarz, dr Alexander Jimenez omawia neurofizjologia kliniczna. Dr Jimenez zbada znaczenie kliniczne i czynności funkcjonalne włókien nerwów obwodowych, rdzenia kręgowego, pnia mózgu i mózgu w kontekście zaburzeń trzewnych i mięśniowo-szkieletowych. Pacjenci uzyskają zaawansowaną wiedzę na temat anatomii, genetyki, biochemii i fizjologii bólu w odniesieniu do różnych zespołów klinicznych. Uwzględniona zostanie biochemia żywieniowa związana z nocycepcją i bólem. A wprowadzenie tych informacji do programów terapeutycznych zostanie podkreślone.
Nasz zespół jest dumny z tego, że dostarcza naszym rodzinom i rannym pacjentom tylko sprawdzone protokoły leczenia. Ucząc pełnego, holistycznego dobrego samopoczucia jako stylu życia, zmieniamy nie tylko życie naszych pacjentów, ale także ich rodzin. Robimy to, abyśmy mogli dotrzeć do jak największej liczby mieszkańców El Paso, którzy nas potrzebują, bez względu na problemy z przystępnością. Aby uzyskać odpowiedzi na wszelkie pytania, zadzwoń do dr Jimenez pod numer 915-850-0900.
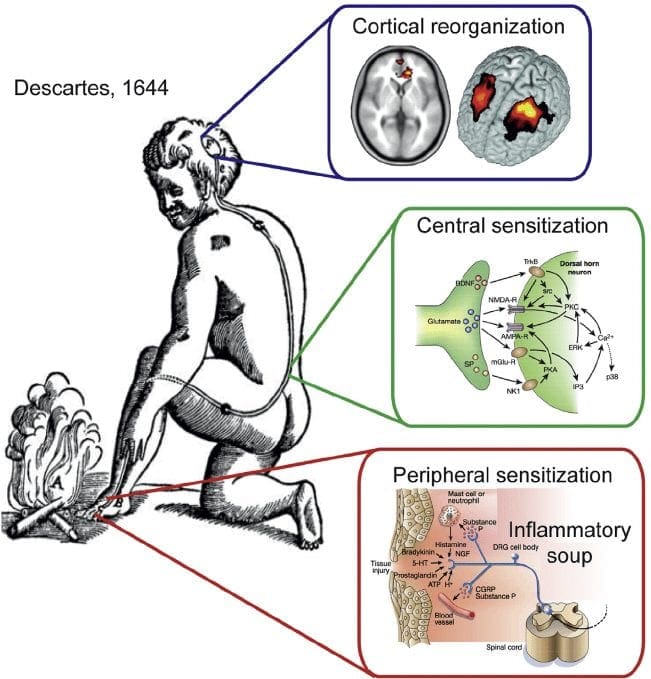
Ból neuropatyczny jest złożonym, przewlekłym stanem bólowym , któremu na ogół towarzyszy uszkodzenie tkanek miękkich. Ból neuropatyczny jest powszechny w praktyce klinicznej i stanowi wyzwanie zarówno dla pacjentów, jak i klinicystów. W przypadku bólu neuropatycznego same włókna nerwowe mogą być uszkodzone, dysfunkcyjne lub uszkodzone. Ból neuropatyczny jest wynikiem uszkodzenia obwodowego lub ośrodkowego układu nerwowego w wyniku urazu lub choroby, gdzie zmiana może wystąpić w dowolnym miejscu. W rezultacie te uszkodzone włókna nerwowe mogą wysyłać nieprawidłowe sygnały do innych ośrodków bólu. Efektem uszkodzenia włókien nerwowych jest zmiana funkcji nerwowej, zarówno w miejscu uszkodzenia, jak i wokół uszkodzenia. Kliniczne objawy bólu neuropatycznego zwykle obejmują zjawiska czuciowe, takie jak ból samoistny, parestezje i przeczulica bólowa.
Ból neuropatyczny, zgodnie z definicją Międzynarodowego Stowarzyszenia Badań Bólu lub IASP, jest bólem zapoczątkowanym lub spowodowanym przez pierwotne uszkodzenie lub dysfunkcję układu nerwowego. Może to wynikać z uszkodzenia w dowolnym miejscu wzdłuż neurosi: obwodowego układu nerwowego, rdzeniowego lub nadrdzeniowego układu nerwowego. Cechy, które odróżniają ból neuropatyczny od innych rodzajów bólu, obejmują ból i objawy czuciowe utrzymujące się poza okresem rekonwalescencji. Charakteryzuje się u ludzi spontanicznym bólem, allodynią lub doświadczaniem nieszkodliwej stymulacji jako bolesnej oraz kauzalgią lub uporczywym palącym bólem. Ból spontaniczny obejmuje uczucie mrowienia, pieczenia, przeszywania, przeszywający i napadowy lub ból podobny do porażenia prądem, często związany z dysestezjami i parestezjami. Wrażenia te zmieniają nie tylko aparat sensoryczny pacjenta, ale także jego samopoczucie, nastrój, uwagę i myślenie. Na ból neuropatyczny składają się zarówno objawy „negatywne”, takie jak utrata czucia i uczucie mrowienia, jak i objawy „dodatnie”, takie jak parestezje, ból samoistny i zwiększone odczuwanie bólu.
Stany często związane z bólem neuropatycznym można podzielić na dwie główne grupy: ból z powodu uszkodzenia ośrodkowego układu nerwowego i ból z powodu uszkodzenia obwodowego układu nerwowego. Do tej pierwszej grupy należą udary korowe i podkorowe, urazowe urazy rdzenia kręgowego, jamistość rdzenia kręgowego i jamistość opuszki, nerwobóle nerwu trójdzielnego i językowo-gardłowego, zmiany nowotworowe i inne zajmujące przestrzeń. Do tej ostatniej grupy należą: ucisk lub uwięźnięcie nerwów, neuropatia niedokrwienna, polineuropatie obwodowe, pleksopatie, ucisk korzeni nerwowych, ból po amputacji kikuta i kończyny fantomowej, neuralgia popółpaścowa i neuropatie nowotworowe.
Patofizjologia bólu neuropatycznego
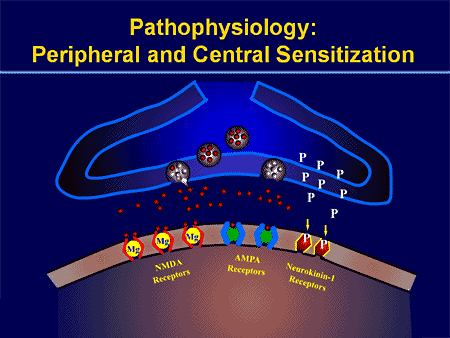
Procesy patofizjologiczne i koncepcje leżące u podstaw bólu neuropatycznego są wielorakie. Przed omówieniem tych procesów krytyczny jest przegląd zwykłych obwodów bólowych. Regularne obwody bólu obejmują aktywację nocyceptora, znanego również jako receptor bólu, w odpowiedzi na bolesną stymulację. Do neuronów pierwszego rzędu dostarczana jest fala depolaryzacji, wraz z wypływem sodu przez kanały sodowe i wypływem potasu. Neurony kończą się w pniu mózgu w jądrze trójdzielnym lub w rogu grzbietowym rdzenia kręgowego. To tutaj znak otwiera bramkowane napięciem kanały wapniowe w terminalu presynaptycznym, umożliwiając wnikanie wapnia. Wapń umożliwia uwalnianie glutaminianu, pobudzającego neuroprzekaźnika do obszaru synaptycznego. Glutaminian wiąże się z receptorami NMDA na neuronach drugiego rzędu, powodując depolaryzację.
Neurony te przechodzą przez rdzeń kręgowy i wędrują aż do wzgórza, gdzie łączą się z neuronami trzeciego rzędu. Te następnie łączą się z układem limbicznym i korą mózgową. Istnieje również ścieżka hamująca, która zapobiega przekazywaniu sygnału bólu z rogu grzbietowego. Neurony antynocyceptywne powstają w pniu mózgu i przemieszczają się w dół rdzenia kręgowego, gdzie łączą się z krótkimi interneuronami w rogu grzbietowym, uwalniając dopaminę i norepinefrynę. Interneurony modulują synapsę między neuronem pierwszego rzędu i neuronem drugiego rzędu poprzez uwalnianie kwasu gamma-aminomasłowego lub GABA, neuroprzekaźnika hamującego. W konsekwencji ustanie bólu jest wynikiem hamowania synaps między neuronami pierwszego i drugiego rzędu, podczas gdy nasilenie bólu może być wynikiem supresji hamujących połączeń synaptycznych.
Mechanizm leżący u podstaw bólu neuropatycznego nie jest jednak tak jasny. Kilka badań na zwierzętach wykazało, że zaangażowanych może być wiele mechanizmów. Trzeba jednak pamiętać, że to, co dotyczy stworzeń, nie zawsze dotyczy ludzi. Neurony pierwszego rzędu mogą zwiększyć swoje odpalanie, jeśli są częściowo uszkodzone i zwiększyć ilość kanałów sodowych. Wyładowania ektopowe są konsekwencją wzmożonej depolaryzacji w niektórych miejscach włókna, powodując ból samoistny i ból związany z ruchem. Obwody hamujące mogą być zmniejszone w poziomie rogu grzbietowego lub komórek macierzystych mózgu, a także obu, co pozwala na swobodny przepływ impulsów bólowych.
Ponadto mogą wystąpić zmiany w centralnym przetwarzaniu bólu, gdy z powodu przewlekłego bólu i stosowania niektórych leków i/lub leków neurony drugiego i trzeciego rzędu mogą tworzyć „pamięć” bólu i ulegać uwrażliwieniu. Następuje wtedy zwiększona wrażliwość neuronów rdzeniowych i zmniejszone progi aktywacji. Inna teoria przedstawia koncepcję bólu neuropatycznego podtrzymywanego współczulnie. Pojęcie to zostało zademonstrowane przez analgezję po sympatektomii od zwierząt i ludzi. Jednak w wielu przewlekłych stanach bólu neuropatycznego lub mieszanych somatycznych i neuropatycznych może być zaangażowana mieszanka mechaniki. Wśród tych wyzwań w dziedzinie bólu, a tym bardziej, jeśli chodzi o ból neuropatyczny, jest możliwość jego sprawdzenia. Jest w tym podwójny składnik: po pierwsze, ocena jakości, intensywności i zaawansowania; a po drugie, prawidłowe diagnozowanie bólu neuropatycznego.
Istnieją jednak pewne narzędzia diagnostyczne, które mogą pomóc klinicystom w ocenie bólu neuropatycznego. Na początek, badania przewodnictwa nerwowego i czuciowych potencjałów wywołanych mogą identyfikować i określać ilościowo zakres uszkodzeń szlaków czuciowych, ale nie nocyceptywnych, poprzez monitorowanie neurofizjologicznych odpowiedzi na bodźce elektryczne. Dodatkowo ilościowe badanie sensoryczne krok po kroku percepcję w reakcji na bodźce zewnętrzne o różnym natężeniu poprzez zastosowanie stymulacji na skórze. Wrażliwość mechaniczna na bodźce dotykowe mierzy się specjalistycznymi narzędziami, takimi jak włosy von Freya, nakłuwanie szpilkami z zazębiającymi się igłami, a także wrażliwość na wibracje wraz z wibrametrami i ból termiczny za pomocą termodów.
Niezwykle ważne jest również przeprowadzenie kompleksowej oceny neurologicznej w celu wykrycia dysfunkcji ruchowych, czuciowych i autonomicznych. Ostatecznie istnieje wiele kwestionariuszy służących do rozróżniania bólu neuropatycznego w bólu nocyceptywnym. Niektóre z nich zawierają tylko pytania do wywiadu (np. kwestionariusz neuropatyczny i ból identyfikacyjny), podczas gdy inne zawierają zarówno pytania do wywiadu, jak i testy fizyczne (np. skala Leeds Assessment of Neuropathic Symptoms and Signs) oraz zupełnie nowe narzędzie, Standaryzowana Ocena Ból, który łączy sześć pytań do wywiadu i dziesięć ocen fizjologicznych.
Metody leczenia bólu neuropatycznego
Schematy farmakologiczne ukierunkowane są na mechanizmy bólu neuropatycznego. Jednak zarówno leczenie farmakologiczne, jak i niefarmakologiczne przynoszą całkowitą lub częściową ulgę tylko u około połowy pacjentów. Wiele opinii opartych na dowodach sugeruje stosowanie mieszanin leków i/lub leków, aby działały z jak największą liczbą mechanizmów. Większość badań dotyczyła głównie neuralgii popółpaścowej i bolesnych neuropatii cukrzycowych, ale wyniki mogą nie dotyczyć wszystkich stanów związanych z bólem neuropatycznym.
Leki przeciwdepresyjne
Leki przeciwdepresyjne zwiększają poziom synaptycznej serotoniny i norepinefryny, wzmacniając w ten sposób działanie zstępującego układu przeciwbólowego związanego z bólem neuropatycznym. Są podstawą terapii bólu neuropatycznego. Działanie przeciwbólowe można przypisać blokadzie wychwytu zwrotnego noradrenaliny i dopaminy, które prawdopodobnie nasilają hamowanie zstępujące, antagonizm receptora NMDA i blokadę kanału sodowego. Trójpierścieniowe leki przeciwdepresyjne, takie jak TCA; np. amitryptylina, imipramina, nortryptylina i doksepina są silne przeciwko ciągłemu bólowi lub palącemu bólowi wraz z bólem spontanicznym.
Udowodniono, że trójpierścieniowe leki przeciwdepresyjne są znacznie bardziej skuteczne w bólu neuropatycznym niż specyficzne inhibitory wychwytu zwrotnego serotoniny lub SSRI, takie jak fluoksetyna, paroksetyna, sertralina i citalopram. Powodem może być to, że hamują one wychwyt zwrotny serotoniny i noradrenaliny, podczas gdy SSRI hamują tylko wychwyt zwrotny serotoniny. Trójpierścieniowe leki przeciwdepresyjne mogą powodować nieprzyjemne skutki uboczne, w tym nudności, splątanie, blokadę przewodzenia w sercu, tachykardię i komorowe zaburzenia rytmu. Mogą również powodować przyrost masy ciała, obniżony próg drgawkowy i niedociśnienie ortostatyczne. Tricykliki należy stosować ostrożnie u osób starszych, które są szczególnie narażone na ich ostre skutki uboczne. Stężenie leku we krwi należy monitorować, aby uniknąć toksyczności u pacjentów wolno metabolizujących leki.
Inhibitory wychwytu zwrotnego serotoniny i noradrenaliny (SNRI) to nowa klasa leków przeciwdepresyjnych. Podobnie jak TCA, wydają się być bardziej skuteczne niż SSRI w leczeniu bólu neuropatycznego, ponieważ hamują również wychwyt zwrotny zarówno noradrenaliny, jak i dopaminy. Wenlafaksyna jest tak samo skuteczna w przypadku wyniszczających polineuropatii, takich jak bolesna neuropatia cukrzycowa, jak imipramina, we wzmiance o TCA, a obie są znacznie silniejsze niż placebo. Podobnie jak TCA, SNRI wydają się przynosić korzyści niezależnie od ich działania przeciwdepresyjnego. Skutki uboczne obejmują uspokojenie polekowe, splątanie, nadciśnienie i zespół odstawienia.
Leki przeciwpadaczkowe
Leki przeciwpadaczkowe mogą być stosowane jako leczenie pierwszego rzutu, zwłaszcza w przypadku niektórych rodzajów bólu neuropatycznego. Działają poprzez modulację kanałów wapniowych i sodowych bramkowanych napięciem, poprawiając hamujące działanie GABA i hamując pobudzającą transmisję glutaminergiczną. Nie wykazano, aby leki przeciwpadaczkowe były skuteczne w ostrym bólu. W przypadkach bólu przewlekłego leki przeciwpadaczkowe wydają się być skuteczne tylko w neuralgii nerwu trójdzielnego. Karbamazepina jest rutynowo stosowana w tym stanie. Wiadomo, że gabapentyna, która działa poprzez hamowanie funkcji kanału wapniowego poprzez działanie agonistyczne w podjednostce alfa-2 delta kanału wapniowego, jest również skuteczna w bólu neuropatycznym. Jednak gabapentyna działa ośrodkowo i może powodować zmęczenie, splątanie i senność.
Nieopioidowe leki przeciwbólowe
Brakuje mocnych danych wspierających stosowanie niesteroidowych leków przeciwzapalnych lub NLPZ w łagodzeniu bólu neuropatycznego. Może to być spowodowane brakiem składnika zapalnego w łagodzeniu bólu. Ale były one stosowane zamiennie z opioidami jako adiuwanty w leczeniu bólu nowotworowego. Odnotowano jednak powikłania, zwłaszcza u pacjentów poważnie osłabionych.
Opioidowe leki przeciwbólowe
Opioidowe środki przeciwbólowe są przedmiotem wielu dyskusji w łagodzeniu bólu neuropatycznego. Działają poprzez hamowanie ośrodkowych wznoszących się impulsów bólowych. Tradycyjnie, ból neuropatyczny był wcześniej obserwowany jako oporny na opioidy, przy czym opioidy są bardziej odpowiednimi metodami w przypadku bólu wieńcowego i somatycznego bólu nocyceptywnego. Wielu lekarzy zapobiega stosowaniu opioidów w leczeniu bólu neuropatycznego, w dużej mierze z powodu obaw związanych z nadużywaniem narkotyków, uzależnieniem i kwestiami regulacyjnymi. Istnieje jednak wiele prób, które wykazały sukces opioidowych środków przeciwbólowych. Oksykodon był lepszy od placebo w łagodzeniu bólu, allodynii, poprawie snu i niepełnosprawności. Opioidy o kontrolowanym uwalnianiu, zgodnie z zaplanowaną podstawą, są zalecane pacjentom z ciągłym bólem w celu pobudzenia stałego poziomu analgezji, zapobiegania wahaniom stężenia glukozy we krwi i zapobiegania zdarzeniom niepożądanym związanym z wyższym dawkowaniem. Najczęściej stosuje się preparaty doustne ze względu na ich większą łatwość stosowania i opłacalność. Preparaty przezskórne, pozajelitowe i doodbytnicze są na ogół stosowane u pacjentów, którzy nie tolerują leków doustnych.
Miejscowe środki znieczulające
Pobliskie środki znieczulające są atrakcyjne, ponieważ dzięki swojemu działaniu regionalnemu mają minimalne skutki uboczne. Działają stabilizując kanały sodowe w aksonach obwodowych neuronów pierwszego rzędu. Działają najlepiej, jeśli występuje tylko częściowe uszkodzenie nerwów i zgromadzony nadmiar kanałów sodowych. Miejscowa lidokaina jest najlepiej przebadanym przedstawicielem kursu bólu neuropatycznego. W szczególności zastosowanie tego 5-procentowego plastra z lidokainą do neuralgii poopryszczkowej spowodowało jej zatwierdzenie przez FDA. Plaster wydaje się działać najlepiej, gdy występuje uszkodzona, ale utrzymana funkcja nocyceptorów obwodowego układu nerwowego z zajętego dermatomu, co przejawia się jako allodynia. Musi być nakładany bezpośrednio na obszar objawowy przez 12 godzin i usuwany przez kolejne 12 godzin i może być stosowany w ten sposób przez lata. Oprócz miejscowych reakcji skórnych jest często dobrze tolerowany przez wielu pacjentów z bólem neuropatycznym.
Różne leki
Wykazano, że klonidyna, alfa-2-agonista, jest skuteczna w podgrupie pacjentów z obwodową neuropatią cukrzycową. Stwierdzono, że kannabinoidy odgrywają rolę w eksperymentalnej modulacji bólu w modelach zwierzęcych i coraz więcej jest dowodów na ich skuteczność. Selektywni agoniści CB2 tłumią hiperalgezję i allodynię oraz normalizują progi nocyceptywne bez wywoływania analgezji.
Interwencyjne leczenie bólu
U pacjentów z nieuleczalnym bólem neuropatycznym można rozważyć leczenie inwazyjne. Zabiegi te obejmują nadtwardówkowe lub okołonerwowe iniekcje miejscowych środków znieczulających lub kortykosteroidów, implantację nadtwardówkowych i dooponowych metod dostarczania leków oraz wprowadzenie stymulatorów rdzenia kręgowego. Podejścia te są zarezerwowane dla pacjentów z nieuleczalnym przewlekłym bólem neuropatycznym, u których nie powiodło się zachowawcze leczenie zachowawcze, a także przeszli gruntowną ocenę psychologiczną. W badaniu Kim i wsp. wykazano, że stymulator rdzenia kręgowego był skuteczny w leczeniu bólu neuropatycznego pochodzenia nerwowego.
Wgląd doktora Alexa Jimeneza
W przypadku bólu neuropatycznego pojawiają się objawy bólu przewlekłego spowodowane uszkodzeniem, dysfunkcją lub uszkodzeniem samych włókien nerwowych, czemu zazwyczaj towarzyszy uszkodzenie lub uraz tkanki. W rezultacie te włókna nerwowe mogą zacząć wysyłać nieprawidłowe sygnały bólu do innych obszarów ciała. Skutki bólu neuropatycznego spowodowanego uszkodzeniami włókien nerwowych obejmują modyfikacje funkcji nerwów zarówno w miejscu uszkodzenia, jak i wokół uszkodzenia. Zrozumienie patofizjologii bólu neuropatycznego było celem wielu pracowników służby zdrowia, aby skutecznie określić najlepsze podejście do leczenia, które pomoże opanować i łagodzić jego objawy. Od stosowania leków i/lub leków, po opiekę chiropraktyczną, ćwiczenia, aktywność fizyczną i odżywianie, można stosować różne metody leczenia, aby pomóc złagodzić ból neuropatyczny zgodnie z potrzebami każdej osoby.
Dodatkowe interwencje w bólu neuropatycznym
Wielu pacjentów z bólem neuropatycznym wybiera uzupełniające i alternatywne opcje leczenia bólu neuropatycznego. Inne dobrze znane schematy stosowane w leczeniu bólu neuropatycznego obejmują akupunkturę, przezskórną elektryczną stymulację nerwów, przezskórną elektryczną stymulację nerwów, leczenie poznawczo-behawioralne, stopniowane obrazowanie motoryczne i leczenie wspomagające oraz ćwiczenia. Wśród nich jednak chiropraktyka jest dobrze znaną alternatywną metodą leczenia, powszechnie stosowaną do pomocy w leczeniu bólu neuropatycznego. Opieka chiropraktyczna wraz z fizjoterapią, ćwiczeniami fizycznymi, odżywianiem i modyfikacjami stylu życia może ostatecznie przynieść ulgę w objawach bólu neuropatycznego.
Chiropraktyka opieki
Wiadomo, że kompleksowa aplikacja do leczenia ma kluczowe znaczenie w zwalczaniu skutków bólu neuropatycznego. W ten sposób chiropraktyka jest holistycznym programem leczenia, który może być skuteczny w zapobieganiu problemom zdrowotnym związanym z uszkodzeniem nerwów. Opieka chiropraktyczna zapewnia pomoc pacjentom z wieloma różnymi schorzeniami, w tym cierpiącymi na ból neuropatyczny. Osoby cierpiące na ból neuropatyczny często stosują niesteroidowe leki przeciwzapalne lub NLPZ, takie jak ibuprofen lub silne leki przeciwbólowe na receptę, aby złagodzić ból neuropatyczny. Mogą one stanowić tymczasowe rozwiązanie, ale wymagają ciągłego używania, aby poradzić sobie z bólem. To niezmiennie przyczynia się do powstawania szkodliwych skutków ubocznych, aw skrajnych sytuacjach uzależnienia od leków na receptę.
Opieka chiropraktyczna może pomóc złagodzić objawy bólu neuropatycznego i zwiększyć stabilność bez tych wad. Podejście takie jak chiropraktyka oferuje zindywidualizowany program mający na celu wskazanie pierwotnej przyczyny problemu. Korzystając z korekty kręgosłupa i manipulacji manualnych, kręgarz może ostrożnie korygować wszelkie niewspółosiowość kręgosłupa lub podwichnięcia występujące wzdłuż kręgosłupa, co może obniżyć konsekwencje niszczenia nerwów poprzez zmianę ustawienia kręgosłupa. Przywrócenie integralności kręgosłupa jest niezbędne do utrzymania sprawnie funkcjonującego ośrodkowego układu nerwowego.
Kręgarz może być również długoterminową terapią mającą na celu poprawę ogólnego samopoczucia. Poza korektami kręgosłupa i manipulacjami manualnymi, kręgarz może zaoferować porady żywieniowe, takie jak przepisanie diety bogatej w przeciwutleniacze lub może opracować fizjoterapię lub program ćwiczeń, aby zwalczyć nawroty bólu nerwowego. Długotrwałe schorzenie wymaga długotrwałego środka zaradczego, a w tym charakterze nieoceniony może być pracownik służby zdrowia specjalizujący się w urazach i/lub schorzeniach układu mięśniowo-szkieletowego i nerwowego, taki jak lekarz kręgarz lub kręgarz. ocenić korzystne zmiany w czasie.
Wykazano, że fizjoterapia, ćwiczenia i techniki reprezentacji ruchu są korzystne w leczeniu bólu neuropatycznego. Opieka chiropraktyczna oferuje również inne metody leczenia, które mogą być pomocne w leczeniu lub poprawie bólu neuropatycznego. Na przykład terapia laserowa niskiego poziomu lub LLLT zyskała ogromne znaczenie w leczeniu bólu neuropatycznego. Zgodnie z różnymi badaniami naukowymi stwierdzono, że LLLT ma pozytywny wpływ na kontrolę znieczulenia bólu neuropatycznego, jednak wymagane są dalsze badania naukowe w celu zdefiniowania protokołów leczenia, które podsumowują efekty terapii laserowej niskiego poziomu w leczeniu bólu neuropatycznego.
Opieka chiropraktyczna obejmuje również porady żywieniowe, które mogą pomóc w kontrolowaniu objawów związanych z neuropatią cukrzycową. Podczas badań naukowych wykazano, że niskotłuszczowa dieta roślinna poprawia kontrolę glikemii u pacjentów z cukrzycą typu 2. Po około 20 tygodniach badania pilotażowego zaangażowane osoby zgłosiły zmiany w masie ciała, a przewodność elektrochemiczna skóry stopy poprawiła się po interwencji. Badania naukowe sugerowały potencjalną wartość interwencji dietetycznej na bazie roślin o niskiej zawartości tłuszczu w neuropatii cukrzycowej. Co więcej, badania kliniczne wykazały, że doustne stosowanie L-treonianu magnezu jest w stanie zapobiegać, a także przywracać deficyty pamięci związane z bólem neuropatycznym.
Opieka chiropraktyczna może również oferować dodatkowe strategie leczenia promujące regenerację nerwów. Na przykład sugerowano, że zwiększenie regeneracji aksonów pomaga poprawić odzyskiwanie funkcji po uszkodzeniu nerwów obwodowych. Stwierdzono, że stymulacja elektryczna, wraz z ćwiczeniami lub aktywnością fizyczną, promuje regenerację nerwów po opóźnionej naprawie nerwów u ludzi i szczurów, zgodnie z ostatnimi badaniami naukowymi. Zarówno stymulacja elektryczna, jak i ćwiczenia zostały ostatecznie uznane za obiecujące eksperymentalne metody leczenia uszkodzenia nerwów obwodowych, które wydają się gotowe do zastosowania klinicznego. Konieczne mogą być dalsze badania naukowe, aby w pełni określić ich skutki u pacjentów z bólem neuropatycznym.
Wnioski
Ból neuropatyczny to wieloaspektowa jednostka, bez szczególnych wytycznych, którymi należy się zająć. Najlepszym sposobem zarządzania jest podejście multidyscyplinarne. Leczenie bólu wymaga ciągłej oceny, edukacji pacjenta, zapewnienia obserwacji i zapewnienia pacjenta. Ból neuropatyczny jest stanem przewlekłym, który stanowi wyzwanie dla najlepszego leczenia. Indywidualizacja leczenia obejmuje rozważenie wpływu bólu na samopoczucie jednostki, depresję i niepełnosprawność wraz z dalszą edukacją i oceną. Badania bólu neuropatycznego, zarówno na poziomie molekularnym, jak i na modelach zwierzęcych, są stosunkowo nowe, ale bardzo obiecujące. Oczekuje się wielu ulepszeń w podstawowych i klinicznych dziedzinach bólu neuropatycznego, co otwiera drogę do ulepszonych lub nowych metod leczenia tego upośledzającego stanu. Zakres naszych informacji ogranicza się do chiropraktyki oraz urazów i stanów kręgosłupa. Aby omówić ten temat, prosimy zapytać dr Jimeneza lub skontaktować się z nami pod adresem�915-850-0900 .
Kurator: dr Alex Jimenez
Dodatkowe tematy: Back Pain
Na ból pleców jest jedną z najczęstszych przyczyn niepełnosprawności i nieobecności w pracy na całym świecie. W rzeczywistości, ból pleców został przypisany jako drugi najczęstszy powód wizyt u lekarza, przewyższający jedynie infekcje górnych dróg oddechowych. Około 80 procent populacji doświadczy pewnego rodzaju bólu pleców przynajmniej raz w ciągu całego życia. Kręgosłup jest złożoną strukturą zbudowaną z kości, stawów, więzadeł i mięśni oraz innych miękkich tkanek. Z tego powodu urazy i / lub pogorszenie warunków, takich jak przepukliny, może ostatecznie prowadzić do objawów bólu pleców. Urazy sportowe lub wypadki samochodowe są często najczęstszą przyczyną bólu pleców, jednak czasami najprostsze ruchy mogą mieć bolesne skutki. Na szczęście alternatywne opcje leczenia, takie jak opieka chiropraktyczna, mogą pomóc złagodzić ból pleców poprzez zastosowanie korekcji kręgosłupa i manualnych manipulacji, ostatecznie poprawiając ulgę w bólu.
Ból Lęk Depresja„Każdy doświadczył bólu, ale są też osoby z depresją, lękiem lub jednym i drugim. Połącz to z bólem i może stać się dość intensywny i trudny do wyleczenia. Osoby cierpiące na depresję, stany lękowe lub jedno i drugie mają tendencję do odczuwania silnego i długotrwałego bólu bardziej niż inni ludzie.
Droga niepokój, depresja i ból nakładają się na siebie jest widoczne w przewlekłych i niektórych zespołach bólowych powodujących niepełnosprawność, tj. bólu krzyża, bóle głowy, nerwoból i fibromialgia. Zaburzenia psychiczne przyczyniają się do nasilenia bólu, a także zwiększają ryzyko niepełnosprawności.
Depresja:A (poważne zaburzenie depresyjne lub depresja kliniczna) jest powszechnym, ale poważnym zaburzeniem nastroju. Powoduje poważne objawy, które wpływają na to, jak dana osoba się czuje, myśli i jak radzi sobie z codziennymi czynnościami, tj. spaniem, jedzeniem i pracą. Aby zdiagnozować depresję, objawy muszą być obecne przez co najmniej dwa tygodnie.
Utrzymujący się smutny, niespokojny lub „pusty” nastrój.
Poczucie beznadziejności, pesymistyczne.
Drażliwość.
Poczucie winy, bezwartościowości lub bezradności.
Utrata zainteresowania lub przyjemności z zajęć.
Zmniejszona energia lub zmęczenie.
Powoli porusza się lub mówi.
Uczucie niepokoju i problemy z siedzeniem w bezruchu.
Trudności z koncentracją, zapamiętywaniem lub podejmowaniem decyzji.
Trudności ze snem, wczesne budzenie się i zasypianie.
Apetyt i zmiany wagi.
Myśli o śmierci lub samobójstwie i lub próbach samobójczych.
Bóle, bóle głowy, skurcze lub problemy z trawieniem bez wyraźnej przyczyny fizycznej i/lub które nie ustępują podczas leczenia.
Nie każdy, kto cierpi na depresję, doświadcza każdego objawu. Niektórzy doświadczają tylko kilku objawów, podczas gdy inni mogą doświadczać kilku. Kilka utrzymujących się objawów oprócz obniżonego nastroju to�wymaganydo diagnozy dużej depresji. Nasilenie i częstotliwość objawów wraz z czasem trwania będą się różnić w zależności od osoby i jej konkretnej choroby. Objawy mogą się również różnić w zależności od stadium choroby.
BÓL LEK DEPRESJA
Cele:
Jaki jest związek?
Jaka za tym stoi neurofizjologia?
Jakie są główne konsekwencje?
Zmiany w mózgu w bólu
Rycina 1 Ścieżki mózgowe, regiony i sieci zaangażowane w ostry i przewlekły ból
Davis, KD i in. (2017) Badania obrazowania mózgu w bólu przewlekłym: kwestie medyczne, prawne i etyczne oraz zalecenia Nat. Wielebny Neurol. doi:10.1038/nrneurol.2017.122
BÓL, LĘK I DEPRESJA
Wnioski:
Ból, zwłaszcza przewlekły, wiąże się z depresją i lękiem
Fizjologiczne mechanizmy prowadzące do lęku i depresji mogą mieć wieloczynnikowy charakter
Kiedy system sensoryczny jest dotknięty urazem lub chorobą, nerwy w tym systemie nie mogą prawidłowo przekazywać wrażeń i uczuć do mózgu. Często przyczynia się to do uczucia drętwienia lub braku czucia. Jednak w niektórych przypadkach, gdy ten system jest uszkodzony, ludzie mogą odczuwać ból w dotkniętym obszarze.
Ból neuropatyczny nie zaczyna się nagle ani nie ustępuje szybko; to jest przewlekły ból stan, który prowadzi do uporczywych objawów bólowych. U większości osób intensywność objawów może nasilać się i słabnąć w ciągu dnia. Chociaż ból neuropatyczny ma być związany z problemami zdrowotnymi nerwów obwodowych, takimi jak neuropatia spowodowana cukrzycą lub zwężeniem kręgosłupa, urazy mózgu lub rdzenia kręgowego mogą również prowadzić do przewlekłego bólu neuropatycznego. Ból neuropatyczny jest również określany jako ból nerwowy.
Ból neuropatyczny można skontrastować z bólem nocyceptywnym. Ból neuropatyczny nie rozwija się do żadnych konkretnych okoliczności ani bodźców zewnętrznych, ale objawy pojawiają się po prostu dlatego, że układ nerwowy może nie działać odpowiednio. W rzeczywistości ludzie mogą również odczuwać ból neuropatyczny, nawet jeśli bolącej lub uszkodzonej części ciała w rzeczywistości nie ma. Ten stan nazywa się bólem kończyn fantomowych, który może wystąpić u osób po amputacji.
Ból nocyceptywny jest na ogół ostry i rozwija się w odpowiedzi na określone okoliczności, na przykład gdy ktoś dozna nagłego urazu, takiego jak walenie w palec młotkiem lub ukłucie palca u stopy podczas chodzenia boso. Co więcej, ból nocyceptywny ma tendencję do ustępowania po wygojeniu się dotkniętego miejsca. Organizm zawiera wyspecjalizowane komórki nerwowe, znane jako nocyceptory, które wykrywają szkodliwe bodźce, które mogą uszkodzić organizm, takie jak ekstremalne ciepło lub zimno, nacisk, szczypanie i narażenie na chemikalia. Te sygnały ostrzegawcze są następnie przekazywane przez układ nerwowy do mózgu, powodując ból nocyceptywny.
Jakie są czynniki ryzyka bólu neuropatycznego?
Wszystko, co przyczynia się do braku funkcji czuciowego układu nerwowego, może prowadzić do bólu neuropatycznego. W związku z tym problemy zdrowotne nerwów związane z zespołem cieśni nadgarstka lub podobnymi stanami mogą ostatecznie wywołać ból neuropatyczny. Uraz, powodujący uszkodzenie nerwów, może prowadzić do bólu neuropatycznego. Inne stany, które mogą predysponować osoby do rozwoju bólu neuropatycznego, obejmują: cukrzycę, niedobory witamin, raka, HIV, udar, stwardnienie rozsiane, półpasiec, a nawet niektóre terapie przeciwnowotworowe.
Jakie są przyczyny bólu neuropatycznego?
Istnieje wiele przyczyn, z których u poszczególnych osób może rozwinąć się ból neuropatyczny. Ale na poziomie komórkowym jednym z wyjaśnień jest zwiększone uwalnianie pewnych receptorów, które wskazują na ból, wraz ze zmniejszoną zdolnością nerwów do modulowania tych sygnałów, co prowadzi do odczuwania bólu pochodzącego z dotkniętego obszaru. Dodatkowo, w rdzeniu kręgowym, obszar bolesny ulega przebudowie z odpowiednimi zmianami hormonalnymi i utratą normalnie funkcjonujących ciał ruchomych. Zmiany te powodują odczuwanie bólu przy braku stymulacji zewnętrznej. W mózgu zdolność do blokowania bólu może zostać zaburzona po urazie, takim jak udar lub uraz po urazie. W miarę upływu czasu dochodzi do dodatkowych uszkodzeń komórek i odczuwania bólu. Ból neuropatyczny jest również związany z cukrzycą, przewlekłym spożywaniem alkoholu, niektórymi nowotworami, niedoborem witaminy B, chorobami, innymi chorobami nerwowymi, toksynami i określonymi lekami.
Jakie są objawy bólu neuropatycznego?
W przeciwieństwie do innych schorzeń neurologicznych identyfikacja bólu neuropatycznego może być trudna. Jednak może być obecnych kilka, jeśli w ogóle, obiektywnych sygnałów. Pracownicy służby zdrowia muszą rozszyfrować i przetłumaczyć zestaw słów, którymi pacjenci opisują swój ból. Pacjenci mogą opisywać swoje objawy jako ostre, tępe, gorące, zimne, wrażliwe, swędzące, głębokie, szczypiące, piekące, wśród wielu innych terminów opisowych. Dodatkowo niektórzy pacjenci mogą odczuwać ból poprzez lekki dotyk lub nacisk.
Aby pomóc określić, ile bólu mogą odczuwać pacjenci, często stosuje się różne skale. Pacjenci proszeni są o ocenę bólu według wizualnej skali lub wykresu numerycznego. Istnieje wiele przykładów skal bólu, takich jak ta pokazana poniżej. Często zdjęcia twarzy przedstawiające różne poziomy bólu mogą być pomocne, gdy dana osoba ma trudności z opisaniem odczuwanego bólu.
Przewlekły ból a zdrowie psychiczne
Dla wielu wpływ przewlekłego bólu może nie ograniczać się do samego bólu; może również negatywnie wpływać na ich stan psychiczny. Nowe badania naukowe przeprowadzone przez naukowców z Northwestern University w Chicago mogą wyjaśnić, dlaczego osoby cierpiące na przewlekły ból cierpią również na pozornie niepowiązane problemy zdrowotne, takie jak depresja, stres, brak snu i trudności z koncentracją.
Ocena wykazała, że osoby z przewlekłym bólem wykazują różne obszary mózgu, które są zawsze aktywne, w szczególności obszar związany z nastrojem i uwagą. To ciągłe działanie przywraca połączenia nerwowe z mózgu i naraża osoby cierpiące na przewlekły ból na większe ryzyko problemów psychologicznych. Naukowcy zasugerowali, że ciągłe otrzymywanie sygnałów bólu może spowodować zmianę mentalną, która niekorzystnie wpływa na umysł. Przeprogramowanie zmusza ich mózgi do innego poświęcania zasobów umysłowych na wykonywanie codziennych zadań, od matematyki, przez przywoływanie listy zakupów, po uczucie szczęścia.
Związek między bólem a mózgiem został dobrze udokumentowany, przynajmniej anegdotycznie, a wielu pracowników służby zdrowia twierdzi, że na własne oczy widziało, jak stan psychiczny pacjenta może się pogorszyć, gdy znosi przewlekły ból. Błędne wyobrażenia na temat związku ból-mózg mogą wynikać z braku dowodów na to, że ból ma mierzalny, trwały wpływ na mózg. Naukowcy spodziewają się, że dzięki dodatkowym badaniom nad mechanizmami, w jaki sposób przewlekły ból czyni ludzi bardziej podatnymi na zaburzenia nastroju, ludzie będą mogli lepiej zarządzać swoim ogólnym samopoczuciem.
Kultura i przewlekły ból
Wiele rzeczy składa się na sposób, w jaki doświadczamy i wyrażamy ból, jednak ostatnio badacze zasugerowali również, że kultura odnosi się bezpośrednio do wyrażania bólu. Nasze wychowanie i wartości społeczne wpływają na to, jak wyrażamy ból, a także na jego własną naturę, intensywność i długość. Jednak te zmienne nie są tak oczywiste, jak wartości społeczno-psychologiczne, takie jak wiek i płeć.
Badania wskazują, że ból przewlekły jest procesem wieloaspektowym, a współzależność między patofizjologią, czynnikami poznawczymi, afektywnymi, behawioralnymi i społeczno-kulturowymi sprowadza się do tego, co określa się mianem doświadczania bólu przewlekłego. Okazało się, że ból przewlekły jest różnie doświadczany u pacjentów z różnych kultur i narodowości.
Niektóre kultury zachęcają do wyrażania bólu, szczególnie w południowej części Morza Śródziemnego i na Bliskim Wschodzie. Inne osoby tłumią to, jak w wielu lekcjach dla naszych dzieci o odważnym zachowaniu i niepłakaniu. Ból jest rozpoznawany jako część ludzkiego doświadczenia. Jesteśmy skłonni zakładać, że komunikacja o bólu płynnie przekroczy granice kulturowe. Ale ludzie cierpiący podlegają sposobom, w jakie ich cywilizacje nauczyły ich doświadczać i wyrażać ból.
Zarówno osoby cierpiące, jak i pracownicy służby zdrowia mają trudności z komunikowaniem bólu ponad granicami etnicznymi. W sprawach takich jak ból, gdzie skuteczna komunikacja może mieć daleko idące konsekwencje dla opieki medycznej, jakości życia i potencjalnie przeżycia, rola kultury w komunikowaniu bólu pozostaje niedoceniana. Ból uporczywy jest wielowymiarowym, złożonym spotkaniem utworzonym przez przeplatanie się i współwpływ czynników biologicznych i psychospołecznych. Znajomość kulminacji tych czynników ma kluczowe znaczenie dla zrozumienia różnic w ich manifestacji i zarządzaniu.
Jak diagnozuje się ból neuropatyczny?
Rozpoznanie bólu neuropatycznego opiera się na dodatkowej ocenie historii pacjenta. W przypadku podejrzenia uszkodzenia nerwów, analiza nerwów wraz z badaniem może być uzasadniona. Najczęstszym sposobem oceny, czy nerw jest uszkodzony, jest zastosowanie medycyny elektrodiagnostycznej. Ta podspecjalizacja medyczna wykorzystuje techniki badania przewodnictwa nerwowego za pomocą elektromielografii (NCS/EMG). Ocena kliniczna może wykazywać oznaki utraty pracy i może obejmować ocenę lekkiego dotyku, zdolność do odróżniania bólu ostrego od tępego oraz zdolność rozróżniania temperatury, a także ocenę wibracji.
Po dokładnym badaniu klinicznym można zaplanować analizę elektrodiagnostyczną. Badania te są prowadzone przez specjalnie przeszkolonych neurologów i fizjologów. Jeśli podejrzewa się neuropatię, należy przeprowadzić poszukiwanie odwracalnych przyczyn. Może to obejmować funkcję krwi pod kątem niedoborów witamin lub problemów z tarczycą oraz badania obrazowe, aby wykluczyć zmiany strukturalne mające wpływ na rdzeń kręgowy. W zależności od wyników tego badania, może istnieć sposób na zmniejszenie intensywności neuropatii i prawdopodobnie zmniejszenie bólu, którego doświadcza pacjent.
Niestety, w wielu przypadkach nawet dobra kontrola podstawowej przyczyny neuropatii nie może odwrócić bólu neuropatycznego. Jest to często obserwowane u pacjentów z neuropatią cukrzycową. W rzadkich przypadkach mogą wystąpić oznaki zmian w skórze i wzorcu wzrostu włosów w dotkniętym regionie. Te zmiany mogą być związane ze zmianami potu. Zmiany te, jeśli są obecne, mogą pomóc w identyfikacji prawdopodobnej obecności bólu neuropatycznego związanego ze stanem znanym jako zespół złożonego bólu regionalnego.
Wgląd doktora Alexa Jimeneza
Ból neuropatyczny to przewlekły stan bólowy, który na ogół wiąże się z bezpośrednim uszkodzeniem lub uszkodzeniem układu nerwowego lub nerwów. Ten rodzaj bólu różni się od bólu nocyceptywnego lub typowego odczuwania bólu. Ból nocyceptywny to ostre lub nagłe odczucie bólu, które powoduje, że układ nerwowy wysyła sygnały bólu natychmiast po wystąpieniu urazu. Jednak w przypadku bólu neuropatycznego pacjenci mogą odczuwać przeszywający, palący ból bez bezpośredniego uszkodzenia lub urazu. Zrozumienie możliwych przyczyn bólu neuropatycznego pacjenta w porównaniu z jakimkolwiek innym rodzajem bólu może pomóc pracownikom służby zdrowia znaleźć lepsze sposoby leczenia przewlekłych stanów bólowych.
Jakie jest leczenie bólu neuropatycznego?
W leczeniu bólu neuropatycznego stosuje się różne leki. Większość tych leków jest stosowana poza wskazaniami rejestracyjnymi, co oznacza, że lek został zatwierdzony przez FDA do leczenia różnych schorzeń, a następnie został uznany za korzystny w leczeniu bólu neuropatycznego. Trójpierścieniowe leki przeciwdepresyjne, takie jak amitryptylina, nortryptylina i dezypramina, są od kilku lat przepisywane w leczeniu bólu neuropatycznego.
Niektóre osoby uważają, że mogą one bardzo skutecznie dawać im ulgę. Wykazano, że inne rodzaje leków przeciwdepresyjnych przynoszą ulgę. U niektórych pacjentów stosowano selektywne inhibitory wychwytu zwrotnego serotoniny lub SSRI, takie jak paroksetyna i citalopram oraz inne leki przeciwdepresyjne, takie jak wenlafaksyna i bupropion. Innym częstym leczeniem bólu neuropatycznego są leki przeciwpadaczkowe, w tym karbamazepina, fenytoina, gabapentyna, lamotrygina i inne.
W ostrych przypadkach bolesnej neuropatii, które nie reagują na pośredników pierwszego rzutu, leki zwykle stosowane w leczeniu arytmii serca mogą przynieść pewne korzyści; jednak mogą one prowadzić do znacznych skutków ubocznych i często muszą być ściśle monitorowane. Leki nakładane bezpośrednio na skórę mogą przynieść niektórym pacjentom niewielkie lub odczuwalne korzyści. Powszechnie stosowane postacie obejmują lidokainę (w postaci plastra lub żelu) lub kapsaicynę.
Leczenie bólu neuropatycznego zależy od przyczyny. Jeśli przyczyna jest odwracalna, nerwy obwodowe mogą się zregenerować, a ból ustąpi; niemniej jednak zmniejszenie bólu może potrwać od kilku miesięcy do lat. Kilka innych alternatywnych opcji leczenia, w tym chiropraktyka i fizjoterapia, może być również wykorzystanych w celu złagodzenia napięcia i stresu wzdłuż nerwów, ostatecznie pomagając złagodzić bolesne objawy.
Jakie są prognozy dla bólu neuropatycznego?
Wiele osób z bólem neuropatycznym jest w stanie uzyskać pewną pomoc, nawet jeśli ich ból nie ustępuje. Chociaż ból neuropatyczny nie jest niebezpieczny dla pacjenta, obecność bólu przewlekłego może negatywnie wpływać na jakość życia. Pacjenci z przewlekłym bólem nerwowym mogą cierpieć z powodu braku snu lub zaburzeń nastroju, w tym depresji, lęku i stresu, jak wspomniano powyżej. Z powodu nieodłącznego łysienia i braku czuciowej informacji zwrotnej, pacjenci są narażeni na ryzyko urazu lub infekcji lub nieświadomego spowodowania eskalacji obecnego urazu. Dlatego ważne jest, aby natychmiast zwrócić się o pomoc medyczną i postępować zgodnie ze szczegółowymi wytycznymi skierowanymi przez pracownika służby zdrowia w zakresie bezpieczeństwa i ostrożności.
Czy można zapobiegać bólowi neuropatycznemu?
Najlepszym sposobem zapobiegania bólowi neuropatycznemu jest unikanie rozwoju lub progresji neuropatii. Monitorowanie i zmiana opcji stylu życia, w tym ograniczanie spożycia alkoholu i tytoniu; utrzymywanie prawidłowej wagi w celu zmniejszenia ryzyka cukrzycy, choroby zwyrodnieniowej stawów lub udaru; a posiadanie świetnej ergonomicznej formy w pracy lub podczas uprawiania hobby w celu zmniejszenia ryzyka urazów spowodowanych powtarzającym się stresem to strategie zmniejszające prawdopodobieństwo rozwoju neuropatii i potencjalnego bólu neuropatycznego. W przypadku jakichkolwiek objawów związanych z bólem neuropatycznym należy niezwłocznie zwrócić się o pomoc medyczną, aby zastosować najbardziej odpowiednie podejście do leczenia. Zakres naszych informacji ogranicza się do chiropraktyki oraz urazów i schorzeń kręgosłupa. Aby omówić ten temat, prosimy zapytać dr Jimeneza lub skontaktować się z nami pod adresem�915-850-0900 .
Kurator: dr Alex Jimenez
Dodatkowe tematy: Back Pain
Na ból pleców jest jedną z najczęstszych przyczyn niepełnosprawności i nieobecności w pracy na całym świecie. W rzeczywistości, ból pleców został przypisany jako drugi najczęstszy powód wizyt u lekarza, przewyższający jedynie infekcje górnych dróg oddechowych. Około 80 procent populacji doświadczy pewnego rodzaju bólu pleców przynajmniej raz w ciągu całego życia. Kręgosłup jest złożoną strukturą zbudowaną z kości, stawów, więzadeł i mięśni oraz innych miękkich tkanek. Z tego powodu urazy i / lub pogorszenie warunków, takich jak przepukliny, może ostatecznie prowadzić do objawów bólu pleców. Urazy sportowe lub wypadki samochodowe są często najczęstszą przyczyną bólu pleców, jednak czasami najprostsze ruchy mogą mieć bolesne skutki. Na szczęście alternatywne opcje leczenia, takie jak opieka chiropraktyczna, mogą pomóc złagodzić ból pleców poprzez zastosowanie korekcji kręgosłupa i manualnych manipulacji, ostatecznie poprawiając ulgę w bólu.
Jeśli system czuciowy zostanie dotknięty urazem lub chorobą, nerwy w tym systemie nie mogą przekazywać wrażeń do mózgu. Może to prowadzić do uczucia drętwienia lub braku czucia. W niektórych przypadkach, gdy układ czuciowy jest uszkodzony, osoby mogą odczuwać ból w dotkniętym obszarze. Ból neuropatyczny nie pojawia się szybko lub szybko się kończy. Jest to choroba przewlekła, która prowadzi do „objawów” uporczywy ból. W przypadku wielu osób nasilenie objawów może pojawiać się i znikać w ciągu dnia. Uważa się, że ból neuropatyczny jest związany z problemami nerwów obwodowych, tj. neuropatia spowodowana cukrzycą, zwężeniem kręgosłupa, uszkodzeniem mózgu lub rdzenia kręgowego może również prowadzić do przewlekłego bólu neuropatycznego.
BÓL NEUROPATYCZNY
Cele:
Co to jest?
Jaka jest za tym patofizjologia?
Jakie są przyczyny
Jakie są niektóre ścieżki?
Jak możemy to naprawić?
BÓL NEUROPATYCZNY
Ból wywołany lub spowodowany pierwotną zmianą lub dysfunkcją w somatosensorycznym układzie nerwowym.
Ból neuropatyczny jest zwykle przewlekły, trudny do leczenia i często oporny na standardowe leczenie przeciwbólowe.
PATOGENEZA BÓLU NEUROPATYCZNEGO
MECHANIZMY PERYFERYJNE
Po uszkodzeniu nerwów obwodowych, neurony stają się bardziej wrażliwe i rozwijają nienaturalną pobudliwość i podwyższoną wrażliwość na stymulację
Jest to znane jako ... Uczulenie peryferyjne!
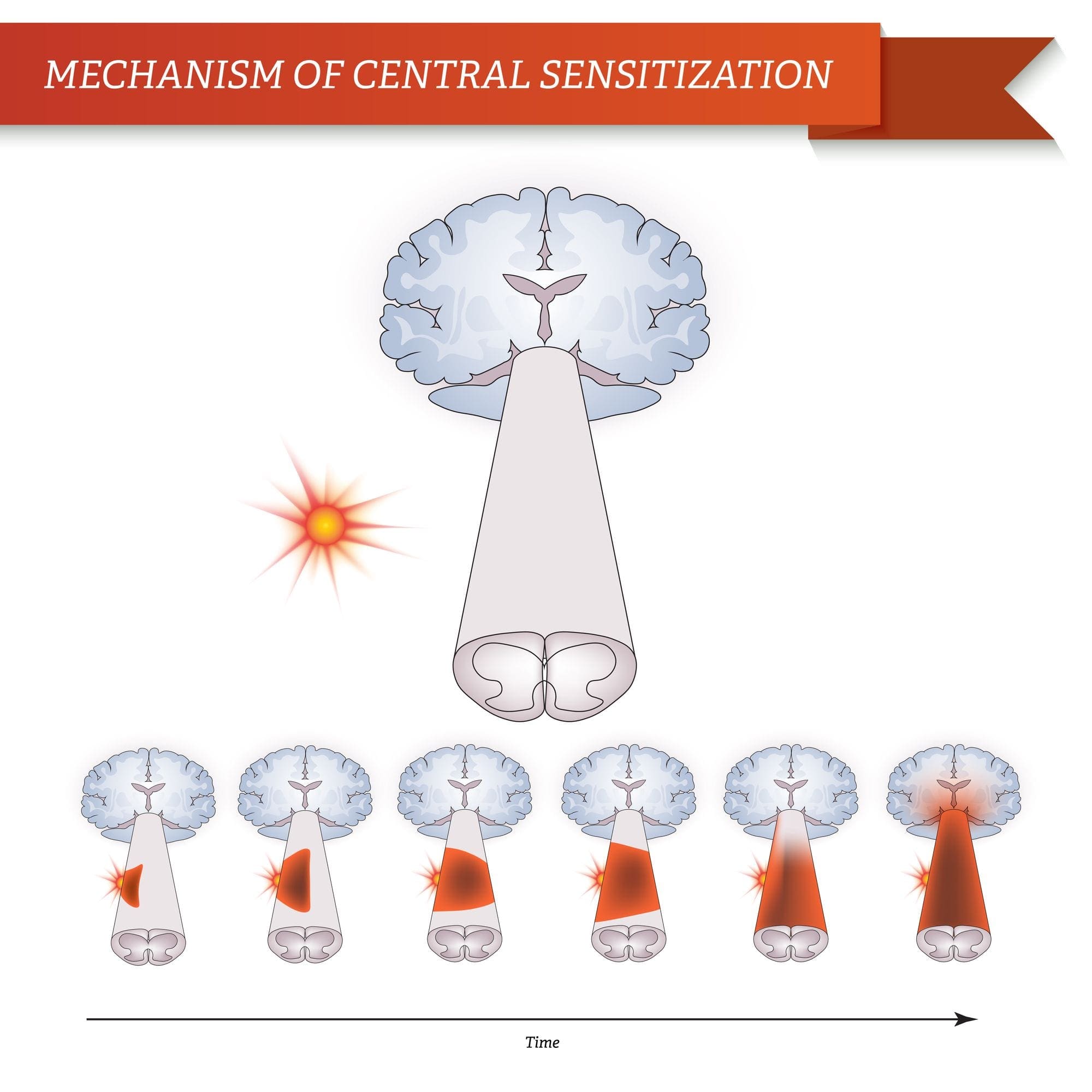
MECHANIZMY CENTRALNE
W wyniku trwającej spontanicznej aktywności na peryferiach, neurony rozwijają zwiększoną aktywność tła, powiększone pola receptywne i zwiększoną odpowiedź na impulsy aferentne, w tym normalne bodźce dotykowe
Jest to znane jako…Uczulenie centralne!
NAJCZĘSTSZE PRZYCZYNY
Uszkodzenia lub choroby somatosensorycznego układu nerwowego mogą prowadzić do zmienionej i zaburzonej transmisji sygnałów czuciowych do rdzenia kręgowego i mózgu; typowe stany związane z bólem neuropatycznym obejmują:
Nerwoból poopryszczkowy
Neuralgia nerwu trójdzielnego
Bolesna radikulopatia
Neuropatia cukrzycowa
Zakażenie wirusem HIV
Trąd
Amputacja
Ból po urazie nerwów obwodowych
Udar (w postaci centralnego bólu poudarowego)
BÓL PHANTOMOWY I RZECZYWISTOŚĆ ROZSZERZONA
Ból kończyn fantomowych i AR
ZAPALENIE NEUROGENNE
Cele:
Co to jest?
Jaka jest za tym patofizjologia?
Jakie są przyczyny
Jak możemy to naprawić?
ZAPALENIE NEUROGENNE
Zapalenie neurogenne jest nerwowo wywołaną, miejscową reakcją zapalną charakteryzującą się rozszerzeniem naczyń krwionośnych, zwiększoną przepuszczalnością naczyń, degranulacją komórek tucznych i uwalnianiem neuropeptydów, w tym SP i peptydu związanego z genem kalcytoniny (CGRP)
Wydaje się, że odgrywa ważną rolę w patogenezie wielu chorób, w tym migreny, łuszczycy, astmy, fibromialgii, egzemy, trądziku różowatego, dystonii i nadwrażliwości chemicznej
NAJCZĘSTSZE PRZYCZYNY
Istnieje wiele dróg, którymi można zainicjować zapalenie neurogenne. Jest dobrze udokumentowane, przy użyciu zarówno modeli zwierzęcych, jak i izolowanych neuronów in vitro, że kapsaicyna, ciepło, protony, bradykinina i tryptaza są regulatorami w górę do wewnątrzkomórkowego napływu wapnia, co powoduje uwalnianie neuropeptydu zapalnego. Przeciwnie, uważa się, że prostaglandyny E2 i I2, cytokiny, interleukina-1, interleukina-6 i czynnik martwicy nowotworu same nie powodują uwalniania neuroprzekaźników, ale raczej pobudzają neurony czuciowe, a tym samym obniżają próg wyzwalania i powodują zwiększone uwalnianie neuropeptydy.
Podczas gdy zapalenie neurogenne zostało szeroko zbadane i dobrze udokumentowane w tkankach obwodowych, do niedawna koncepcja zapalenia neurogennego w OUN pozostawała w dużej mierze niezbadana. Biorąc pod uwagę zdolność neurogennego zapalenia do wpływania na przepuszczalność naczyń i do powstawania obrzęku, obecnie jest szeroko badany pod kątem jego potencjalnego wpływu na przepuszczalność BBB i obrzęk naczyniopochodny w mózgu i rdzeniu kręgowym w różnych stanach patologicznych.
Wszystkich nas w dzieciństwie uczono, że istnieje 5 zmysłów: wzrok, smak, dźwięk, węch i dotyk. Początkowe cztery zmysły wykorzystują jasne, odrębne narządy, takie jak oczy, kubki smakowe, uszy i nos, ale jak dokładnie ciało odczuwa dotyk? Dotyk odczuwany jest na całym ciele, zarówno wewnątrz, jak i na zewnątrz. Nie ma jednego odrębnego organu odpowiedzialnego za wyczuwanie dotyku. Wokół całego ciała znajdują się raczej maleńkie receptory lub zakończenia nerwowe, które wyczuwają dotyk w miejscu jego wystąpienia i wysyłają do mózgu sygnały z informacjami dotyczącymi rodzaju dotyku, który miał miejsce. Ponieważ kubek smakowy na języku wykrywa smak, mechanoreceptory to gruczoły znajdujące się w skórze i na innych narządach, które wykrywają wrażenia dotykowe. Są znani jako mechanoreceptory ponieważ są zaprojektowane do wykrywania mechanicznych odczuć lub różnic w ciśnieniu.
Rola mechanoreceptorów
Osoba rozumie, że doświadczyła wrażenia, gdy organ odpowiedzialny za odkrycie tego konkretnego zmysłu wysyła wiadomość do mózgu, który jest głównym organem przetwarzającym i porządkującym wszystkie informacje. Wiadomości są wysyłane ze wszystkich obszarów ciała do mózgu za pośrednictwem przewodów zwanych neuronami. Istnieją tysiące małych neuronów, które rozgałęziają się na wszystkie obszary ludzkiego ciała, a na zakończeniach wielu z tych neuronów znajdują się mechanoreceptory. Aby pokazać, co się dzieje, gdy dotykasz przedmiotu, użyjemy przykładu.
Wyobraź sobie, że komar ląduje na twoim ramieniu. Szczep tego owada, tak lekki, stymuluje mechanoreceptory w tym konkretnym obszarze ramienia. Te mechanoreceptory wysyłają wiadomość wzdłuż neuronu, z którym są połączone. Neuron łączy się przez całą drogę z mózgiem, który otrzymuje wiadomość, że coś dotyka twojego ciała w dokładnej lokalizacji konkretnego mechanoreceptora, który wysłał wiadomość. Mózg będzie działał zgodnie z tą radą. Może powie oczom, żeby spojrzały na obszar ramienia, który wykrył podpis. A kiedy oczy mówią mózgowi, że na ramieniu jest komar, mózg może powiedzieć dłoni, aby szybko go odrzuciła. Tak działają mechanoreceptory. Celem poniższego artykułu jest wykazanie i szczegółowe omówienie organizacji funkcjonalnej i molekularnych uwarunkowań mechanoreceptorów.
Zmysł dotyku: organizacja funkcjonalna i determinanty molekularne mechanowrażliwych receptorów
Abstrakcyjny
Mechanoreceptory skórne są zlokalizowane w różnych warstwach skóry, gdzie wykrywają szeroki zakres bodźców mechanicznych, w tym lekkie szczotkowanie, rozciąganie, wibracje i szkodliwy nacisk. Tej różnorodności bodźców odpowiada różnorodna gama wyspecjalizowanych mechanoreceptorów, które reagują na deformację skóry w określony sposób i przekazują te bodźce do wyższych struktur mózgu. Badania nad mechanoreceptorami i genetycznie wyprowadzalnymi zakończeniami nerwów czuciowych zaczynają odkrywać mechanizmy odczuwania dotyku. Prace w tej dziedzinie umożliwiły naukowcom dokładniejsze zrozumienie organizacji obwodów leżących u podstaw percepcji dotyku. Nowe kanały jonowe pojawiły się jako kandydaci na cząsteczki transdukcji, a właściwości prądów bramkowanych mechanicznie poprawiły naszą wiedzę na temat mechanizmów adaptacji do bodźców dotykowych. W niniejszym przeglądzie podkreślono postępy poczynione w charakteryzowaniu właściwości funkcjonalnych mechanoreceptorów w owłosionej i gładkiej skórze oraz kanałach jonowych, które wykrywają sygnały mechaniczne i adaptację mechanoreceptorów kształtu.
Słowa kluczowe:mechanoreceptor, kanał mechanowrażliwy, ból, skóra, układ somatosensoryczny, dotyk
Wprowadzenie
Dotyk to wykrywanie bodźców mechanicznych oddziałujących na skórę, w tym nieszkodliwych i szkodliwych bodźców mechanicznych. Jest to zasadniczy sens dla przetrwania i rozwoju ssaków i człowieka. Kontakt ciał stałych i płynów ze skórą dostarcza niezbędnych informacji do ośrodkowego układu nerwowego, które umożliwiają eksplorację i rozpoznawanie otoczenia oraz inicjują lokomocję lub zaplanowany ruch ręki. Dotyk jest również bardzo ważny dla nauki zawodu, kontaktów towarzyskich i seksualności. Zmysł dotyku jest najmniej wrażliwym zmysłem, choć może być zniekształcony (przeczulica, hipoestezja) w wielu stanach patologicznych.1-3
Reakcje na dotyk polegają na bardzo precyzyjnym kodowaniu informacji mechanicznych. Mechanoreceptory skóry są zlokalizowane w różnych warstwach skóry, gdzie wykrywają szeroki zakres bodźców mechanicznych, w tym lekkie szczotkowanie, rozciąganie, wibracje, odchylanie włosów i szkodliwy nacisk. Tej różnorodności bodźców odpowiada różnorodna gama wyspecjalizowanych mechanoreceptorów, które reagują na deformację skóry w określony sposób i przekazują te bodźce do wyższych struktur mózgu. Neurony somatosensoryczne skóry dzielą się na dwie grupy: niskoprogowe mechanoreceptory (LTMR) reagujące na łagodny nacisk i wysokoprogowe mechanoreceptory (HTMR) reagujące na szkodliwą stymulację mechaniczną. Ciała komórek LTMR i HTMR znajdują się w zwojach korzeni grzbietowych (DRG) i zwojach czuciowych czaszki (zwojach trójdzielnych). Włókna nerwowe związane z LTMR i HTMR są klasyfikowane jako włókna Aβ-, Aβ- lub C na podstawie ich potencjalnych prędkości przewodzenia. Włókna C są niezmielinizowane i mają najwolniejsze prędkości przewodzenia (~2 m/s), podczas gdy A? i A? włókna są lekko i silnie zmielinizowane, wykazując odpowiednio pośrednią (~12 m/s) i szybką (~20 m/s) prędkość przewodzenia. LTMR są również klasyfikowane jako wolno lub szybko adaptujące się odpowiedzi (SA- i RA-LTMR) na podstawie ich szybkości adaptacji do utrzymującego się bodźca mechanicznego. Ponadto wyróżniają się skórnymi narządami końcowymi, które unerwiają, oraz preferowanymi bodźcami.
Zdolność mechanoreceptorów do wykrywania sygnałów mechanicznych zależy od obecności kanałów jonowych mechanoprzetworników, które szybko przekształcają siły mechaniczne w sygnały elektryczne i depolaryzują pole odbiorcze. Ta lokalna depolaryzacja, zwana potencjałem receptora, może generować potencjały czynnościowe, które rozprzestrzeniają się w kierunku ośrodkowego układu nerwowego. Jednak właściwości cząsteczek, które pośredniczą w mechanotransdukcji i adaptacji do sił mechanicznych, pozostają niejasne.
W tym przeglądzie przedstawiamy przegląd właściwości mechanoreceptorów ssaków w nieszkodliwym i szkodliwym dotyku w skórze owłosionej i gładkiej. Uwzględniamy również najnowszą wiedzę o właściwościach prądów bramkowanych mechanicznie, próbując wyjaśnić mechanizm adaptacji mechanoreceptorów. Na koniec dokonujemy przeglądu ostatnich postępów w identyfikacji kanałów jonowych i powiązanych białek odpowiedzialnych za generowanie prądów bramkowanych mechanicznie.
Niewinny dotyk
LTMR związane z mieszkiem włosowym
Mieszki włosowe reprezentują mini-organy wytwarzające łodygi włosa, które wykrywają lekki dotyk. Włókna związane z mieszkami włosowymi reagują na ruch włosa i jego kierunek, uruchamiając na początku ciągi potencjałów czynnościowych i usuwając bodziec. Szybko przystosowują się do receptorów.
Kot i królik. W sierści kota i królika mieszki włosowe można podzielić na trzy typy mieszków włosowych: włosy puchowe, włosy Guard i Tylotrichs. Najliczniejszymi, najkrótszymi i najdelikatniejszymi włosami sierści są włosy puchowe (podwłos, wełna, welus)4. Są pofalowane, bezbarwne i pojawiają się w grupach od dwóch do czterech włosów ze wspólnego otworu w skórze. Włosy Gwardii (jednokrotne, nadsłuchowe, górne)4 są lekko zakrzywione, pigmentowane lub niepigmentowane i wyłaniają się pojedynczo z ust ich mieszków włosowych. Tylotrichs są najmniej liczne, najdłuższe i najgrubsze.5,6 Są one pigmentowane lub niepigmentowane, czasami jedno i drugie, i wychodzą pojedynczo z mieszka otoczonego pętlą naczyń włosowatych. Włókna czuciowe dostarczane do mieszków włosowych znajdują się poniżej gruczołów łojowych i przypisuje się je A? lub włókna A?-LTMR.7
W bliskim kontakcie z łodygą włosa, tuż poniżej poziomu gruczołu łojowego, znajduje się pierścień lancetowatych zakończeń pilo-ruffini. Te zakończenia nerwów czuciowych są umieszczone spiralnie wokół trzonu włosa w obrębie tkanki łącznej tworzącej mieszek włosowy. W mieszkach włosowych znajdują się również wolne zakończenia nerwowe, niektóre z nich tworzą mechanoreceptory. Często ciałka dotykowe (patrz gładka skóra) otaczają obszar szyi pęcherzyka tylotricha.
Właściwości zmielinizowanych zakończeń nerwowych w owłosionej skórze kota i królika były intensywnie badane w okresie 1930-1970 (przegląd w Hamann, 1995).8 Co ciekawe, Brown i Iggo, badając 772 jednostki z mielinowanymi włóknami doprowadzającymi nerwów odpiszczelowych u kota i królików, sklasyfikowali odpowiedzi w trzech typach receptorów odpowiadających ruchom włosów typu Down (receptory typu D), włosów Guard (receptory typu G) i włosów Tylotrich (receptor typu T). w receptorze szybko adaptacji typu I (RA I) w opozycji do receptora Paciniego o nazwie RA II. Mechanoreceptory RA I wykrywają prędkość bodźca mechanicznego i mają ostrą granicę. Nie wykrywają zmian termicznych. Burgess i in. opisali również szybko przystosowujący się receptor pola, który optymalnie reaguje na głaskanie skóry lub ruch kilku włosów, co przypisano stymulacji zakończeń pilo-Ruffini. Żadna z odpowiedzi mieszków włosowych nie została przypisana aktywności włókien C.9
Myszy. W grzbietowej skórze owłosionej myszy opisano trzy główne typy mieszków włosowych: zygzakowaty (około 72%), szydło/auchene (około 23%) oraz strażnik lub tylotrich (około 5%).11-14 Zygzak i szydło/ Mieszki włosowe auchenne wytwarzają cieńsze i krótsze łodygi włosów i są związane z jednym gruczołem łojowym. Włosy Guard lub Tylotrich są najdłuższymi typami mieszków włosowych. Charakteryzują się dużą cebulką włosową związaną z dwoma gruczołami łojowymi. Włosy ochronne i szydło/auchene są ułożone w powtarzalny, regularnie rozmieszczony wzór, podczas gdy włosy zygzakowate gęsto zasiedlają obszary skóry otaczające dwa większe typy mieszków włosowych [ryc. 1 (A1, A2 i A3)].
Rysunek 1. Organizacja i projekcje mechanoreceptorów skórnych. W przypadku skóry owłosionej lekkie szczotkowanie i dotyk są wykrywane głównie przez unerwienie wokół mieszków włosowych: szydło/auchenne (A1), zygzak (A2) i osłona (A3). Włosy szydło/auchene są potrójnie unerwione przez lancetowate zakończenia C-LTMR (A4), A?-LTMR i A? szybko przystosowujący się-LTMR (A6). Zygzakowate mieszki włosowe są krótszymi włosami i są unerwione zarówno przez C-LTMR (A4), jak i A? -Lancetowate zakończenia LTMR (A5). Najdłuższe mieszki włosowe ochronne są unerwione przez A? szybko przystosowujące się podłużne lancetowate zakończenia LTMR (A6) i są związane z A? powoli adaptujący się – LTMR końcówek kopuł dotykowych (A7). Centralne występy wszystkich tych włókien kończą się odrębnymi, ale częściowo zachodzącymi na siebie blaszkami rogu grzbietowego rdzenia kręgowego (C-LTMR w blaszce II, Ap-LTMR w blaszce III i Ap-LTMR w blaszce IV i V). Projekcje LTMR, które unerwiają te same lub sąsiednie mieszki włosowe, są wyrównane, tworząc wąską kolumnę w rogu grzbietowym rdzenia kręgowego (B1 w kolorze szarym). Jedynie w skórze owłosionej subpopulacja wolnych końcówek włókien C unerwia naskórek i reaguje na przyjemny dotyk (A8). Te włókna C-touch nie reagują na szkodliwy dotyk, a ich droga nie jest jeszcze znana (B2). W przypadku gładkiej skóry w nieszkodliwym dotyku pośredniczą cztery typy LTMR. Kompleks komórka Merkla z neurytem znajduje się w warstwie podstawnej naskórka (C1). Ten mechanoreceptor składa się z układu między wieloma komórkami Merkela i powiększonym zakończeniem nerwu z jednego A? włókno. Komórki Merkla wykazują podobne do palców procesy kontaktu z keratynocytami (C2). Zakończenie Ruffini zlokalizowane jest w skórze właściwej. Jest to cienkie końcówki sensoryczne w kształcie cygara połączone z A? włókno (C3). Ciałka Meissnera połączona z A? zakończenie nerwowe i znajduje się w brodawkach skórnych. Ten zamknięty w kapsułce mechanoreceptor składa się z upakowanych komórek podtrzymujących ułożonych w poziome lamele otoczone tkanką łączną (C4). Ciałko Paciniego jest głębszym mechanoreceptorem. Jedno A? niezmielinizowane zakończenie nerwowe kończy się w środku tej dużej jajowatej ciałki utworzonej z koncentrycznych blaszek. Wypustki tych włókien Aa-LTMR w rdzeniu kręgowym są podzielone na dwie gałęzie. Główna gałąź centralna (B3) wznosi się w rdzeniu kręgowym w ipsilateralnych grzbietowych, tworzących pęczkach klinowych lub smukłych (B5) na poziomie rdzenia, gdzie pierwotne aferenty tworzą swoją pierwszą synapsę (B6). Neurony drugorzędowe tworzą decussation czuciowe (B7), tworząc szlak na przyśrodkowym lemnisku, który wznosi się przez pień mózgu do śródmózgowia, szczególnie we wzgórzu. Druga gałąź LTMR kończy się w rogu grzbietowym w blaszce II, IV, V i zakłóca przenoszenie bólu (B4). Szkodliwy dotyk jest wykrywany przez wolne zakończenie nerwowe w naskórku zarówno skóry owłosionej (A9), jak i gładkiej (C7). Te mechanoreceptory stanowią zakończenie A?-HTMR i C-HTMR w bliskim kontakcie z sąsiednimi keratynocytami (C6). A?-hTMR kończy się w blaszce I i V; C-HTMR kończy się w blaszce I i II (B8). Na poziomie rogu grzbietowego rdzenia kręgowego, pierwotne aferentne HTMR tworzą synapsy z wtórnymi neuronami, które przecinają linię środkową i wspinają się do wyższej struktury mózgu w przednio-bocznym pęczku (B9, B10).
Ostatnio Ginty i współpracownicy zastosowali kombinację metod znakowania molekularno-genetycznego i somatotopowego śledzenia wstecznego, aby zwizualizować organizację obwodowych i centralnych zakończeń aksonów LTMR u myszy.15 Ich odkrycia potwierdzają model, w którym występują indywidualne cechy złożonego bodźca dotykowego. ekstrahowane przez trzy typy mieszków włosowych i przenoszone poprzez działanie unikalnych kombinacji włókien A'-, A'- i C- do rogu grzbietowego.
Wykazali, że genetyczne znakowanie neuronów DRG dodatnich pod względem hydroksylazy tyrozynowej (TH+) charakteryzuje populację niepeptydergicznych neuronów czuciowych o małej średnicy i umożliwia wizualizację zakończeń obwodowych C-LTMR w skórze. Co zaskakujące, odkryto, że gałęzie aksonalne poszczególnych C-LTMR drzewiają się i tworzą podłużne lancetowate zakończenia, które są ściśle związane z zygzakiem (80% zakończeń) i szydłem/auchene (20% zakończeń), ale nie mieszków włosowych tylotrich [Ryc. 1 (A4)]. Od dawna uważano, że podłużne zakończenia lancetowate należą wyłącznie do A?-LTMR i dlatego nieoczekiwane było, że zakończenia C-LTMR będą tworzyć podłużne zakończenia lancetowate. zmielinizowane mechanoreceptory [ryc. 15 (C2)].
Rysunek 2. Receptory dotykowe u ssaków: Receptory dotykowe na skórze różnicują się w nieszkodliwy dotyk, wspierany przez wiele receptorów o niskim progu mechanicznym (LTMR) w skórze gładkiej i owłosionej oraz szkodliwy dotyk wspierany przez receptor o wysokim progu mechanicznym (HTMR). Tworzą beznerwowe zakończenia, które kończą się głównie w naskórku. (A) Skóra gładka. A1: Ciałka Meissnera wykrywają ruch skóry i ślizganie się przedmiotu w dłoni. Są ważne dla przekazywania przedmiotów i zręczności. Receptory szybko przystosowują się do bodźca, czy są połączone z A? włókna i rzadko do włókien C i mają duże pole receptorowe. A2: Ciałka Ruffiniego wykrywają rozciągnięcie skóry i są ważne, aby wykryć pozycję palców i przedmiot, który podaje. Receptor powoli adaptuje się do bodźca i utrzymuje aktywność tak długo, jak bodziec był stosowany. Receptory są podłączone do A? włókna i mają duże pole odbioru. O3: Ciałka Paciniego znajdują się głębiej w skórze właściwej i wykrywają wibracje. Receptory są podłączone do A? włókna; szybko przystosowują się do bodźca i mają największe pole odbioru. (B) Cała skóra. B1: Kompleksy komórek Merkla są obecne zarówno w gładkiej skórze, jak i wokół włosów. Są one gęsto wyrażone w dłoni i są ważne dla percepcji tekstury i najlepszego rozróżniania między dwoma punktami. Odpowiadają za precyzję palców. Receptory są podłączone do A? włókna; powoli przystosowują się do bodźca i mają krótkie pole recepcyjne. B2: Szkodliwy dotyk HTMR z bardzo powolną adaptacją do bodźca, tj. aktywny tak długo, jak długo stosowany jest bodziec nocyceptywny. Tworzą je wolne zakończenia nerwowe A? i włókna C związane z keratynocytami. (C) Owłosiona skóra. C1: Mieszki włosowe są powiązane z różnymi typami włosów. U myszy włosy Guard są dłuższe i słabo wyeksponowane, szydło/auchenne są średniej wielkości, a zygzaki są najmniejszymi i najgęściej wyeksponowanymi włosami. Są połączone z A? włókna, ale także do A? oraz włókna C-LTMRs do włosów szydło/auchenne i zyzag. Wykrywają ruch włosów, w tym przyjemny dotyk podczas pieszczoty. Przystosowują się do bodźca szybko lub z pośrednią kinetycznością. C2: Zakończenia nerwowe C-touch odpowiadają podtypowi zakończeń włókien C z wolnym zakończeniem charakteryzującym się niskim progiem mechanicznym. Mają kodować przyjemne doznania wywołane pieszczotą. Umiarkowanie przystosowują się do bodźca i mają krótkie pole odbioru. Domniemane mechanowrażliwe (MS) kanały jonowe wyrażane w różnych receptorach dotykowych są wskazywane zgodnie ze wstępnymi danymi i podsumowują obecną hipotezę w trakcie oceny.
Druga główna zidentyfikowana populacja dotyczy zakończeń Ap-LTMR w mieszkach Awl/Auchenne i zygzakowatych w porównaniu z mieszkami włosowymi Downa szeroko badanymi u kotów i królików. Ginty i współpracownicy wykazali, że TrkB ulega ekspresji na wysokim poziomie w podzbiorze neuronów DRG o średniej średnicy. Rejestracje wewnątrzkomórkowe przy użyciu preparacji skórno-nerwowej znakowanych włókien ex vivo wykazały, że wykazują one właściwości fizjologiczne włókien wcześniej badanych u kotów i królików: znakomita wrażliwość mechaniczna (próg von Freya < 0.07 mN), szybko adaptujące się reakcje na bodźce ponadprogowe, przewodzenie pośrednie prędkości (5.8 – 0.9 m/s) i wąskie, nieodkształcone kolce somy.15 Te Aβ-LTMR tworzą podłużne, lancetowate zakończenia związane z praktycznie każdym zygzakowatym i szydłem/ouchene mieszkiem włosowym tułowia [Ryc. 1 (A5)].
Wreszcie wykazali, że peryferyjne zakończenia szybko adaptującego się A? LTMR tworzą podłużne lancetowate zakończenia związane z mieszkami włosowymi osłony (lub tylotrich) i szydła/auchene [Ryc. 1 (A6)].15 Ponadto, włosy Guard są również związane z kompleksem komórek Merkla tworzącym kopułę dotykową połączoną z A? powoli przystosowujący się LTMR [ryc. 1 (A7)].
Podsumowując, praktycznie wszystkie zygzakowate mieszki włosowe są unerwione przez lancetowate zakończenia C-LTMR i Ap-LTMR; Włosy szydła/auchene są potrójnie unerwione przez A? szybko adaptujące się lancetowate zakończenia LTMR, Ap-LTMR i C-LTMR; Ochronne mieszki włosowe są unerwione przez A? szybko dostosowujące się podłużne lancetowate zakończenia LTMR i oddziałują z A? powoli dostosowujące się - LTMR końcówek kopuł dotykowych. W ten sposób każdy mieszek włosowy myszy otrzymuje unikalne i niezmienne kombinacje zakończeń LTMR odpowiadających neurofizjologicznie odmiennym mechanosensorycznym narządom końcowym. Biorąc pod uwagę iteracyjne ułożenie tych trzech rodzajów włosów, Ginty i współpracownicy proponują, aby skóra owłosiona składała się z powtarzających się powtórzeń jednostki peryferyjnej zawierającej (1) jeden lub dwa centralnie umieszczone włosy ochronne, (2) ~20 otaczających włosów szydła/auchenne i (3 ) ~80 przeplatanych zygzakowatych włosków [ryc. 2 (C1)].
Projekcja rdzenia kręgowego. Centralne projekcje A? szybko adaptujące się LTMR, Ap-LTMR i C-LTMR kończą się odrębnymi, ale częściowo zachodzącymi na siebie blaszkami (II, III, IV) rogu grzbietowego rdzenia kręgowego. Ponadto środkowe końcówki LTMR, które unerwiają te same lub sąsiednie mieszki włosowe w obwodowej jednostce LTMR, są wyrównane, tworząc wąską kolumnę LTMR w rogu grzbietowym rdzenia kręgowego [ryc. 1 (B1)]. Wydaje się zatem prawdopodobne, że klin lub kolumna somatotopowo zorganizowanych pierwotnych dośrodkowych zakończeń czuciowych w rogu grzbietowym reprezentuje wyrównanie środkowych wypustek A?-, A?- i C-LTMR, które unerwiają tę samą jednostkę obwodową i wykrywają mechaniczne bodźce działające na tę samą małą grupę mieszków włosowych. Na podstawie liczby włosków ochronnych, szydłowatych i zygzakowatych na tułowiu i kończynach oraz liczby każdego podtypu LTMR, Ginty i współpracownicy szacują, że róg grzbietowy myszy zawiera 2,000-4,000 kolumn LTMR, co odpowiada przybliżonej liczbie obwodowych Jednostki LTMR.15
Co więcej, aksony podtypów LTMR są ze sobą ściśle powiązane, mają splecione wypustki i splecione, lancetowate zakończenia, które unerwiają ten sam mieszek włosowy. Ponadto, ponieważ te trzy typy mieszków włosowych wykazują różne kształty, rozmiary i składy komórkowe, prawdopodobnie będą miały różne właściwości odchylania lub dostrajania wibracji. Te odkrycia są zgodne z klasycznymi pomiarami neurofizjologicznymi u kota i królika wskazującymi, że A? RA-LTMR i Aβ-LTMR mogą być aktywowane w różny sposób przez odchylanie różnych typów mieszków włosowych.16,17
Podsumowując, dotyk w skórze owłosionej jest kombinacją: (1) względnych liczb, unikalnych rozkładów przestrzennych oraz odrębnych właściwości morfologicznych i odchylania trzech typów mieszków włosowych; (2) unikalne kombinacje zakończeń podtypu LTMR związane z każdym z trzech typów mieszków włosowych; oraz (3) odrębne wrażliwości, prędkości przewodzenia, wzorce skoków i właściwości adaptacyjne czterech głównych klas LTMR związanych z mieszkiem włosowym, które umożliwiają systemowi mechanosensorycznemu skóry owłosionej wyodrębnienie i przekazanie do OUN złożonych kombinacji cech, które definiują dotykać.
LTMR zakończeń wolnego nerwu
Ogólnie rzecz biorąc, wolne zakończenia włókien C w skórze to HTMR, ale subpopulacja włókien C nie reaguje na szkodliwy dotyk. Ten podzbiór dośrodkowych włókien dotykowych C (CT) reprezentuje odrębny typ niezmielinizowanych, niskoprogowych jednostek mechanoreceptywnych istniejących w owłosionej, ale nie gładkiej skórze ludzi i ssaków [Fig. 1 (A8)].18,19 CT są ogólnie związane z odczuwaniem przyjemnej stymulacji dotykowej w kontakcie cielesnym.20,21
Aferenty CT reagują na siły wgniatające w zakresie 0.3–2.5 mN, a zatem są tak samo wrażliwe na deformację skóry jak wiele komórek A? aferenty.19 Charakterystyki adaptacyjne neuronów aferentnych CT są zatem pośrednie w porównaniu z wolno i szybko adaptującymi się zmielinizowanymi mechanoreceptorami. Pola recepcyjne ludzkich aferentnych CT mają z grubsza okrągły lub owalny kształt. Pole składa się z jednego do dziewięciu małych reagujących punktów rozmieszczonych na obszarze do 35 mm2.22 Receptory mysich homologów są zorganizowane we wzór nieciągłych łat pokrywających około 50% obszaru skóry owłosionej [ryc. 60 (C2)].2
Dowody pochodzące od pacjentów bez mielinowych włókien aferentnych wskazują, że sygnalizacja we włóknach CT aktywuje korę wyspy. Ponieważ system ten słabo koduje rozróżniające aspekty dotyku, ale dobrze nadaje się do kodowania powolnego, delikatnego dotyku, włókna CT w skórze owłosionej mogą być częścią systemu przetwarzania przyjemnych i społecznie istotnych aspektów dotyku.24 Aktywacja włókien CT może również odgrywają rolę w hamowaniu bólu, a ostatnio zaproponowano, że stan zapalny lub uraz może zmienić odczucie przekazywane przez LTMR z włókna C z przyjemnego dotyku na ból.25,26
Która ścieżka podróżują aferenty CT nie jest jeszcze znana [Ryc. 1 (B2)], ale udokumentowano niskoprogowe sygnały dotykowe do komórek projekcji rdzeniowo-wzgórzowej,27 co potwierdza doniesienia o subtelnych, kontralateralnych deficytach wykrywania dotyku u pacjentów po zniszczeniu tych dróg po zabiegach chordotomii.28
LTMR w Glabrous Skin
Kompleksy komórkowo-neurytowe Merkla i kopułka dotykowa. Merkel (1875) jako pierwsza podała histologiczny opis skupisk komórek naskórka z dużymi zrazionymi jądrami, stykających się z przypuszczalnie doprowadzającymi włóknami nerwowymi. Przypuszczał, że służyły one zmysłowi dotyku, nazywając je Tastzellen (komórki dotykowe). U ludzi kompleksy komórek Merkel neurytów są wzbogacone w wrażliwe na dotyk obszary skóry, znajdują się w podstawowej warstwie naskórka palców, ust i narządów płciowych. Występują również w skórze owłosionej o mniejszej gęstości. Kompleks komórka Merkla – neuryt składa się z komórki Merkla w bliskim kontakcie z powiększonym zakończeniem nerwowym z pojedynczej mielinizowanej A? włókno [ryc. 1 (C1)] (przegląd w Halata i wsp.).29 Po stronie naskórka komórka Merkla wykazuje wyrostki palcowe rozciągające się między sąsiednimi keratynocytami [ryc. 1 (C2)]. Komórki Merkla są komórkami naskórka pochodzącymi z keratynocytów.30,31 Termin kopuła dotykowa została wprowadzona, aby nazwać duże stężenie kompleksów komórek Merkla w owłosionej skórze przedniej łapy kota. Kopuła dotykowa może zawierać do 150 komórek Merkla unerwionych przez pojedyncze włókno A?, au ludzi oprócz włókien A?, A? i włókna C były również regularnie obecne.32-34
Stymulacja kompleksów komórek Merkel-neurytów powoduje powolne dostosowywanie się odpowiedzi typu I (SA I), które pochodzą z punktowych pól receptywnych o ostrych granicach. Nie ma spontanicznego wyładowania. Kompleksy te reagują na głębokość wgłębienia skóry i mają najwyższą rozdzielczość przestrzenną (0.5 mm) spośród mechanoreceptorów skóry. Przekazują one precyzyjny przestrzenny obraz bodźców dotykowych i mają być odpowiedzialne za rozróżnianie kształtu i tekstury [ryc. 2 (B1)]. Myszy pozbawione komórek Merkla nie mogą wykrywać teksturowanych powierzchni stopami, podczas gdy robią to za pomocą wąsów.35
To, czy komórka Merkel, neuron czuciowy, czy oba są miejscami mechanotransdukcji, jest nadal przedmiotem dyskusji. U szczurów fototoksyczne niszczenie komórek Merkel znosi odpowiedź SA I.36 U myszy z genetycznie supresją komórek Merkel reakcja SA I zarejestrowana w preparacie skóry/nerwu ex vivo całkowicie zniknęła, co pokazuje, że komórki Merkel są wymagane do prawidłowego kodowania Merkel. odpowiedzi receptora.37 Jednak mechaniczna stymulacja izolowanych komórek Merkla w hodowli za pomocą nacisku napędzanego motorycznie nie generuje prądów bramkowanych mechanicznie.38,39 Keratynocyty mogą odgrywać ważną rolę w prawidłowym funkcjonowaniu kompleksu komórek Merkla–neurytów. Procesy przypominające palce komórki Merkla mogą poruszać się wraz z deformacją skóry i ruchem komórek naskórka, co może być pierwszym etapem mechanicznej transdukcji. Oczywiście warunki wymagane do badania wrażliwości mechanicznej ogniw Merkel nie zostały jeszcze ustalone.
Końcówki Ruffiniego. Końcówki Ruffini to cienkie końcówki sensoryczne w kształcie cygara połączone z A? zakończenia nerwowe. Zakończenia Ruffini to małe cylindry tkanki łącznej ułożone wzdłuż pasm kolagenu skórnego, które są dostarczane przez od jednego do trzech zmielinizowanych włókien nerwowych o średnicy 4 µm. Maksymalnie trzy cylindry o różnej orientacji w skórze właściwej mogą łączyć się, tworząc jeden receptor [Fig. 6 (C1)]. Strukturalnie zakończenia Ruffini są podobne do ścięgien aparatu Golgiego. Są one szeroko wyrażane w skórze właściwej i zostały zidentyfikowane jako wolno adaptujące się mechanoreceptory skórne typu II (SA II). Na tle spontanicznej aktywności nerwowej powoli adaptujące się regularne wyładowanie jest wywoływane przez prostopadłą niską siłę utrzymywaną mechaniczną stymulację lub bardziej efektywnie przez rozciąganie skóry. Odpowiedź SA II pochodzi z dużych pól receptywnych o niejasnych granicach. Receptory Ruffiniego przyczyniają się do percepcji kierunku ruchu obiektu poprzez wzór rozciągnięcia skóry [Rys. 3 (A2)].
U myszy, odpowiedzi SA I i SA II można oddzielić elektrofizjologicznie w preparacie ex-vivo nerwu do skóry.40 Nandasena i współpracownicy donieśli o immunolokalizacji akwaporyny 1 (AQP1) w zakończeniu przyzębia Ruffini siekaczy szczurów, co sugeruje, że AQP1 bierze udział w utrzymanie równowagi osmotycznej zębów niezbędnej do mechanotransdukcji.41 Zakończenia przyzębia Ruffini wyrażają również przypuszczalny mechanowrażliwy kanał jonowy ASIC3.42
Ciałka Meissnera. Ciałka Meissnera zlokalizowane są w brodawkach naskórka, głównie dłoni i podeszew stóp, ale także warg, języka, twarzy, sutków i genitaliów. Anatomicznie składają się z zamkniętego zakończenia nerwowego, którego kapsułka składa się ze spłaszczonych komórek podporowych ułożonych w poziome blaszki osadzone w tkance łącznej. Jest jedno włókno nerwowe A? aferenty połączone na ciałko [ryc. 1 (C4)]. Każda fizyczna deformacja ciałka wyzwala serię potencjałów czynnościowych, które szybko zanikają, tj. szybko adaptują się one do receptorów. Gdy bodziec zostanie usunięty, ciałko odzyskuje swój kształt, a przy tym wytwarza kolejną salwę potencjałów czynnościowych. Ze względu na powierzchowne umiejscowienie w skórze właściwej ciałka te selektywnie reagują na ruchy skóry, dotykową detekcję poślizgu i wibracje (20-40 Hz). Są wrażliwe na dynamiczną skórę – na przykład między skórą a przedmiotem, który jest obsługiwany [ryc. 2 (A1)].
Ciałka Paciniego. Ciałka Paciniego są głębszymi mechanoreceptorami skóry i są najbardziej wrażliwym zamkniętym mechanoreceptorem ruchu skóry. Te duże jajowate ciałka (1 mm długości) zbudowane z koncentrycznych blaszek włóknistej tkanki łącznej i fibroblastów wyłożonych płaskimi zmodyfikowanymi komórkami Schwanna ulegają ekspresji w głębokiej skórze właściwej43. W centrum ciałka, w wypełnionej płynem jamie zwanej wewnętrzną bańką , kończy jeden pojedynczy A? doprowadzające niezmielinizowane zakończenie nerwowe [ryc. 1 (C5)]. Posiadają duże pole recepcyjne na powierzchni skóry ze szczególnie wrażliwym środkiem. Rozwój i funkcja kilku szybko adaptujących się typów mechanoreceptorów są zaburzone u myszy z mutantem c-Maf. Zwłaszcza ciałka Paciniego ulegają poważnej atrofii44
Ciałka Paciniego wykazują bardzo szybką adaptację w odpowiedzi na wgłębienie skóry, szybko adaptujące się wyładowanie nerwowe II (RA II), które jest zdolne do podążania za wysoką częstotliwością bodźców wibracyjnych i umożliwia percepcję odległych zdarzeń poprzez przekazywane wibracje.45 Ciałka Paciniego aferenty reagują na utrzymujące się wcięcie z przejściową aktywnością na początku i odsunięciu bodźca. Nazywa się je również detektorami przyspieszenia, ponieważ mogą wykrywać zmiany w sile bodźca, a jeśli tempo zmian bodźca ulegnie zmianie (jak to ma miejsce w przypadku wibracji), ich reakcja staje się proporcjonalna do tej zmiany. Ciałka Paciniego wyczuwają duże zmiany ciśnienia, a przede wszystkim drgania (150 Hz), które są w stanie wykryć nawet z odległości centymetra [Rys. 300 (A2)].
Odpowiedź toniczną zaobserwowano w pozbawionych kapsułek ciałkach Paciniego.46 Ponadto, nienaruszone ciałka Paciniego odpowiadają utrzymującą się aktywnością podczas ciągłych bodźców powodujących wgniecenie, bez zmiany progów mechanicznych lub częstotliwości odpowiedzi, gdy sygnalizacja za pośrednictwem GABA jest zablokowana między glejem blaszkowym a zakończeniem nerwu.47 nieneuronowe składniki ciałka Paciniego mogą odgrywać podwójną rolę w filtrowaniu bodźca mechanicznego, jak również w modulowaniu właściwości odpowiedzi neuronu czuciowego.
Projekcje rdzenia kręgowego. Projekcje Aa-LTMR w rdzeniu kręgowym są podzielone na dwie gałęzie. Główna gałąź centralna wznosi się w rdzeniu kręgowym w ipsilateralnych kolumnach grzbietowych do poziomu szyjki macicy [ryc. 1 (B3)]. Gałęzie wtórne kończą się w rogu grzbietowym w blaszce IV i zakłócają na przykład przenoszenie bólu. Może to złagodzić ból jako część kontroli bramy [ryc. 1 (B4)].48
Na poziomie szyjnym aksony głównej gałęzi rozdzielają się na dwa pasy: pas pośrodkowy zawiera pęczek spłaszczony, przenoszący informacje z dolnej połowy ciała (nogi i tułów), a pas zewnętrzny zawiera pęczek klinowy, przenoszący informacje z górnej połowy ciała. ciała (ramiona i tułów) [ryc. 1 (B5)].
Pierwotne włókna aferentne dotykowe tworzą swoją pierwszą synapsę z neuronami drugiego rzędu w rdzeniu, gdzie włókna z każdego odcinka synapsy w jądrze o tej samej nazwie: synapsy aksonów pęczka smukłego w jądrze smukłym i aksonów klinowych w jądrze klinowym [ryc. 1 (B6)]. Neurony otrzymujące synapsę dostarczają wtórne aferenty i natychmiast przecinają linię środkową, tworząc po przeciwnej stronie pnia mózgu – przyśrodkowy lemniscus – który wznosi się przez pień mózgu do następnej stacji przekaźnikowej w śródmózgowiu, a konkretnie we wzgórzu [Ryc. . 1 (B7)].
Molekularna specyfikacja LTMR. Mechanizmy molekularne kontrolujące wczesną dywersyfikację LTMR zostały ostatnio częściowo wyjaśnione. Bourane i współpracownicy wykazali, że populacje neuronów eksprymujące receptor kinazy tyrozynowej Ret (Ret) i jego koreceptor GFR?2 u embrionalnych myszy DRG E11 selektywnie eksprymują czynnik transkrypcyjny Mafa.13 Autorzy ci wykazują, że Mafa/ Neurony Ret/GFRy, które mają stać się po urodzeniu trzema specyficznymi typami LTRM: neurony SA49,50 unerwiające kompleksy komórek Merkla, szybko adaptujące się neurony unerwiające ciałka Meissnera i szybko adaptujące się aferenty (RA I) tworzące lancetowate zakończenia wokół mieszków włosowych. Ginty i współpracownicy donoszą również, że neurony DRG z ekspresją wczesnego Ret szybko dostosowują mechanoreceptory z ciałek Meissnera, ciałek Paciniego i lancetowatych zakończeń wokół mieszków włosowych. projekcje aksonalne neuronów w pniu mózgu.
Badanie mechanoreceptorów skóry człowieka. Technika „mikroneurografii” opisana przez Hagbartha i Vallbo w 1968 r. została zastosowana do badania zachowania wyładowań pojedynczych mechanowrażliwych zakończeń zaopatrujących mięśnie, stawy i skórę (patrz przegląd Macefield, 2005). W badaniach scharakteryzowano fizjologię dotykowych włókien doprowadzających w gładkiej skórze dłoni. Zapisy mikroelektrodowe z nerwu pośrodkowego i łokciowego u ludzi ujawniły wrażenia dotykowe generowane przez cztery klasy LTMR: aferenty Meissnera są szczególnie wrażliwe na przesuwanie się światła po skórze, reagując na lokalne siły ścinające i początkowe lub jawne poślizgi w polu odbioru. Pacinian aferentny są niezmiernie wrażliwe na energiczne mechaniczne transjenty. Aferenty reagują energicznie na dmuchanie nad polem receptywnym. Ciałko Paciniego znajdujące się w palcu zwykle reaguje na stukanie w stół podtrzymujący ramię. Doprowadzające Merkel charakteryzują się wysoką dynamiczną wrażliwością na bodźce wgniatane na dyskretny obszar i często reagują wyładowaniem podczas uwalniania. Chociaż aferenty Ruffiniego reagują na siły normalnie działające na skórę, unikalną cechą aferentnych SA II jest ich zdolność do reagowania również na boczne rozciąganie skóry. Wreszcie, jednostki włosowe na przedramieniu mają duże, jajowate lub nieregularne pola receptywne złożone z wielu wrażliwych punktów, które odpowiadają poszczególnym włosom (każde doprowadzające dostarcza ~52,53 włosów).
Mechaniczna wrażliwość keratynocytów
Każdy bodziec mechaniczny na skórze musi być przenoszony przez keratynocyty tworzące naskórek. Te wszechobecne komórki mogą pełnić funkcje sygnalizacyjne oprócz swoich funkcji wspierających lub ochronnych. Na przykład keratynocyty wydzielają ATP, ważną czuciową cząsteczkę sygnalizacyjną, w odpowiedzi na bodźce mechaniczne i osmotyczne.54,55 Uwalnianie ATP indukuje wewnątrzkomórkowy wzrost wapnia poprzez autokrynną stymulację receptorów purynergicznych.55 Ponadto istnieją dowody, że hipotoniczność aktywuje Rho - szlak sygnałowy kinazy i późniejsze tworzenie włókien stresowych F-aktyny, co sugeruje, że mechaniczne odkształcenie keratynocytów może mechanicznie zakłócać sąsiednie komórki, takie jak komórki Merkela w przypadku nieszkodliwego dotyku i zakończeń wolnych od włókien C w przypadku szkodliwego dotyku [Fig. 1 (C6)].56,57
Trujący dotyk
Mechanoreceptory wysokiego progu (HTMR) to naskórkowe C- i A? darmowe zakończenia nerwowe. Nie są one związane z wyspecjalizowanymi strukturami i są obserwowane zarówno w skórze owłosionej [Ryc. 1 (A9)] i gładką skórę [Ryc. 1(C7)]. Jednak termin „wolne zakończenia nerwowe” należy rozpatrywać rozważnie, ponieważ zakończenia nerwowe są zawsze w bliskim kontakcie z keratynocytem lub komórką Langheransa lub melanocytami. Analiza ultrastrukturalna zakończeń nerwowych wykazała obecność szorstkiej retikulum endoplazmatycznego, obfitych mitochondriów i pęcherzyków o gęstym rdzeniu. Sąsiednie błony komórek naskórka są pogrubione i przypominają błonę postsynaptyczną w tkankach nerwowych. Należy zauważyć, że interakcje między zakończeniami nerwowymi a komórkami naskórka mogą być dwukierunkowe, ponieważ komórki naskórka mogą uwalniać mediatory takie jak ATP, interleukina (IL6, IL10) i bradykinina i odwrotnie, peptydowe zakończenia nerwowe mogą uwalniać peptydy, takie jak CGRP lub substancja P działająca na komórki naskórka. HTMR obejmują mechano-nocyceptory wzbudzane tylko przez szkodliwe bodźce mechaniczne i polimodalne nocyceptory, które również reagują na szkodliwe ciepło i egzogenną substancję chemiczną [Fig. 2 (B2)].58
Włókna doprowadzające HTMR kończą się na neuronach projekcyjnych w rogu grzbietowym rdzenia kręgowego. A?-HTMRs kontaktują się z neuronami drugiego rzędu głównie w blaszce I i V, podczas gdy C-HTMR kończą się w blaszce II [Fig. 1 (B8)]. Neurony nocyceptywne drugiego rzędu rzutują na stronę kontrolną rdzenia kręgowego i wznoszą się w istocie białej, tworząc układ przednio-boczny. Te neurony kończą się głównie we wzgórzu [ryc. 1 (B9 i B10)].
Mechano-prądy w neuronach somatosensorycznych
Mechanizmy powolnej lub szybkiej adaptacji mechanoreceptorów nie zostały jeszcze wyjaśnione. Nie jest jasne, w jakim stopniu adaptację mechanoreceptorów zapewnia środowisko komórkowe zakończenia nerwu czuciowego, wewnętrzne właściwości kanałów bramkowanych mechanicznie oraz właściwości kanałów jonowych bramkowanych napięciem aksonów w neuronach czuciowych (ryc. 2). Jednak ostatnie postępy w charakterystyce prądów bramkowanych mechanicznie wykazały, że w neuronach DRG istnieją różne klasy kanałów mechanowrażliwych i mogą one wyjaśniać niektóre aspekty adaptacji mechanoreceptorów.
Rejestracja in vitro u gryzoni wykazała, że soma neuronów DRG jest wewnętrznie wrażliwa mechanicznie i wyraża prądy kationowo bramkowane. i benzamil powodują częściową blokadę.59 FM64-60,62,63 działa jako trwały bloker, a wstrzyknięcie FM1-43 w tylną łapę myszy zmniejsza wrażliwość na ból w teście Randalla-Selitto i zwiększa oceniany próg wycofania łapy z włosami von Frey.1
W odpowiedzi na ciągłą stymulację mechaniczną, prądy mechanowrażliwe zmniejszają się poprzez zamknięcie. Na podstawie stałych czasowych zaniku prądu wyróżniono cztery różne typy prądów mechanowrażliwych: prądy szybko adaptujące się (~3 ms), prądy pośrednio adaptujące (~6 ms), prądy wolno adaptujące się (~15 30 ms). ) i bardzo wolno adaptujące się prądy (~200 ms).300 Wszystkie te prądy występują ze zmienną częstotliwością w szczurzych neuronach DRG unerwiających gładką skórę tylnej łapy.1000
Czułość mechaniczną prądów mechanowrażliwych można określić, stosując serię przyrostowych bodźców mechanicznych, co pozwala na stosunkowo szczegółową analizę bodziec-prąd.66 Zależność między bodźcem a prądem jest zwykle sigmoidalna, a maksymalna amplituda prądu jest określana przez liczbę kanały, które są jednocześnie otwarte.64,67 Co ciekawe, doniesiono, że szybko adaptujący się prąd mechanowrażliwy wykazuje niski próg mechaniczny i punkt środkowy aktywacji w połowie w porównaniu z bardzo wolno adaptującym się prądem mechanowrażliwym.63,65
Neurony czuciowe o fenotypach nienocyceptywnych preferencyjnie wyrażają szybko adaptujące się prądy mechanowrażliwe z niższym progiem mechanicznym.60,61,63,64,68 Z drugiej strony, wolno i bardzo wolno adaptujące się prądy mechanowrażliwe są sporadycznie zgłaszane w domniemanych komórkach nienocyceptywnych.64,68 To skłoniło do sugestii, że prądy te mogą przyczyniać się do różnych mechanicznych progów obserwowanych w LTMR i HTMR in vivo. Chociaż te eksperymenty in vitro należy podchodzić z ostrożnością, potwierdzenie obecności w somie neuronów DRG nisko- i wysokoprogowych mechanoprzekaźników zapewniła również stymulacja oparta na promieniowym rozciąganiu hodowanych mysich neuronów czuciowych.69 Ten paradygmat ujawnił dwa główne populacje neuronów wrażliwych na rozciąganie, jedna, która odpowiada na niską amplitudę bodźca, a druga, która selektywnie reaguje na wysoką amplitudę bodźca.
Wyniki te mają ważne, ale spekulacyjne implikacje mechanistyczne: mechaniczny próg neuronów czuciowych może mieć niewiele wspólnego z komórkową organizacją mechanoreceptora, ale może leżeć we właściwościach mechanicznie bramkowanych kanałów jonowych.
Mechanizmy leżące u podstaw odczulania mechanowrażliwych prądów kationowych w neuronach DRG szczura zostały niedawno wyjaśnione.64,67 Wynika to z dwóch współbieżnych mechanizmów wpływających na właściwości kanałów: adaptacji i inaktywacji. Adaptację po raz pierwszy opisano w badaniach nad komórkami słuchowymi. Można to opisać operacyjnie jako proste przesunięcie krzywej aktywacji kanału przetwornika wzdłuż osi bodźca mechanicznego.70-72 Adaptacja pozwala receptorom czuciowym na zachowanie wrażliwości na nowe bodźce w obecności bodźca istniejącego. Jednak znaczna część prądów mechanowrażliwych w neuronach DRG nie może zostać reaktywowana po kondycjonowaniu mechanicznej stymulacji, co wskazuje na dezaktywację niektórych kanałów przetwornika.64,67 Zatem zarówno inaktywacja, jak i adaptacja działają równolegle w celu regulacji prądów mechanowrażliwych. Te dwa mechanizmy są wspólne dla wszystkich prądów mechanowrażliwych zidentyfikowanych w neuronach DRG szczura, co sugeruje, że powiązane elementy fizykochemiczne determinują kinetykę tych kanałów.64
Podsumowując, określenie właściwości endogennych prądów mechanowrażliwych in vitro ma kluczowe znaczenie w dążeniu do identyfikacji mechanizmów transdukcji na poziomie molekularnym. Obserwowana zmienność progu mechanicznego i adaptacyjna kinetyka różnych mechanicznie bramkowanych prądów w neuronach DRG sugerują, że wewnętrzne właściwości kanałów jonowych mogą przynajmniej częściowo wyjaśniać mechaniczny próg i kinetykę adaptacji mechanoreceptorów opisane w latach 1960. 80 stosując preparaty ex vivo.
Domniemane białka mechanowrażliwe
Mechanowrażliwe prądy jonowe w neuronach somatosensorycznych są dobrze scharakteryzowane, natomiast niewiele wiadomo na temat tożsamości cząsteczek, które pośredniczą w mechanotransdukcji u ssaków. Badania genetyczne u Drosophila i C. elegans zidentyfikowały potencjalne cząsteczki mechanotransdukcji, w tym rodziny TRP i degenerynowy/nabłonkowy kanał Na+ (Deg/ENaC). 73 Ostatnie próby wyjaśnienia molekularnych podstaw mechanotransdukcji u ssaków w dużej mierze koncentrowały się na homologach tych kandydatów . Ponadto wielu z tych kandydatów jest obecnych w skórnych mechanoreceptorach i neuronach somatosensorycznych (ryc. 2).
Kanały jonowe wykrywające kwas
ASIC należą do podgrupy z bramkowanymi protonami rodziny kanałów Na+ z degeneracją nabłonka.74 Trzech członków rodziny ASIC (ASIC1, ASIC2 i ASIC3) ulega ekspresji w mechanoreceptorach i nocyceptorach. Rola kanałów ASIC została zbadana w badaniach behawioralnych na myszach z celowaną delecją genów kanałów ASIC. Delecja ASIC1 nie zmienia funkcji skórnych mechanoreceptorów, ale zwiększa mechaniczną czułość aferentnych unerwiających jelito.75 Myszy z nokautem ASIC2 wykazują zmniejszoną czułość szybko adaptujących się skórnych LTMR.76 Jednak późniejsze badania wykazały brak wpływu nokautu ASIC2 na zarówno mechanonocycepcja trzewna, jak i mechanosensacja skórna.77 Zakłócenie ASIC3 zmniejsza wrażliwość mechaniczną trzewnych włókien aferentnych i zmniejsza reakcje skórnych HTMR na szkodliwe bodźce.76
Kanał receptora przejściowego
Nadrodzina TRP jest podzielona na sześć podrodzin u ssaków.78 Prawie wszystkie podrodziny TRP mają członków powiązanych z mechanosensacją w różnych układach komórkowych.79 Jednak w neuronach czuciowych ssaków kanały TRP są najlepiej znane z wykrywania informacji termicznych i pośredniczenia w zapaleniu neurogennym. i tylko dwa kanały TRP, TRPV4 i TRPA1, zostały zaangażowane w reakcję na dotyk. Zakłócenie ekspresji TRPV4 u myszy ma jedynie niewielki wpływ na ostre progi mechanosensoryczne, ale silnie zmniejsza wrażliwość na szkodliwe bodźce mechaniczne.80,81 TRPV4 jest kluczowym wyznacznikiem w kształtowaniu odpowiedzi neuronów nocyceptywnych na stres osmotyczny i mechaniczną hiperalgezję podczas zapalenia.82,83, 1 TRPA1 wydaje się odgrywać rolę w przeczulicy mechanicznej. Myszy z niedoborem TRPA1 wykazują nadwrażliwość na ból. TRPA84,85 przyczynia się do transdukcji bodźców mechanicznych, zimnych i chemicznych w nocyceptorowych neuronach czuciowych, ale wydaje się, że nie jest to istotne dla transdukcji komórek włoskowatych.XNUMX
Nie ma jednoznacznych dowodów wskazujących na to, że kanały TRP i kanały ASIC wyrażane u ssaków są bramkowane mechanicznie. Żaden z tych kanałów wyrażonych heterologicznie nie odzwierciedla sygnatury elektrycznej prądów mechanowrażliwych obserwowanych w ich naturalnym środowisku. Nie wyklucza to możliwości, że kanały ASIC i TRP są mechanoprzekaźnikami, biorąc pod uwagę niepewność, czy kanał mechanotransdukcji może funkcjonować poza kontekstem komórkowym (patrz rozdział dotyczący SLP3).
Białka piezoelektryczne
Białka piezoelektryczne zostały niedawno zidentyfikowane przez Coste'a i współpracowników jako obiecujący kandydaci do mechanizacji białek.86,87 Kręgowce mają dwóch członków Piezo, Piezo 1 i Piezo 2, wcześniej znanych odpowiednio jako FAM38A i FAM38B, które są dobrze zachowane u wielokomórkowych eukariontów. . Piezo 2 obfituje w grupy DRG, podczas gdy Piezo 1 jest ledwo wykrywalny. Prądy mechanowrażliwe indukowane przez piezo są hamowane przez gadolin, czerwień rutenową i GsMTx4 (toksyna z ptasznika Grammostola spatulata).88 Ekspresja Piezo 1 lub Piezo 2 w układach heterologicznych wytwarza prądy mechanowrażliwe, a kinetyka inaktywacji prądu Piezo 2 jest szybsza niż piezo 1. Podobnie jak prądy endogenne mechanowrażliwe, prądy piezo-zależne mają potencjały odwrotne około 0 mV i nie są kationowo selektywne, a wszystkie Na+, K+, Ca2+ i Mg2+ przenikają przez leżący poniżej kanał. Podobnie prądy piezozależne są regulowane przez potencjał błonowy, z wyraźnym spowolnieniem kinetyki prądu przy potencjałach zdepolaryzowanych.86
Białka piezo są niewątpliwie białkami mechanoczującymi i mają wiele wspólnych właściwości szybko adaptujących się prądów mechanowrażliwych w neuronach czuciowych. Traktowanie hodowanych neuronów DRG krótkim interferującym RNA Piezo 2 zmniejszyło odsetek neuronów z szybko adaptującym się prądem i zmniejszył odsetek neuronów mechanowrażliwych.86 Domeny transbłonowe są zlokalizowane w białkach piezoelektrycznych, ale nie stwierdzono wyraźnych motywów zawierających pory ani sygnatur kanałów jonowych zidentyfikowane. Jednak mysie białko Piezo 1 oczyszczone i zrekonstytuowane w asymetryczne dwuwarstwy lipidowe i liposom tworzy kanały jonowe wrażliwe na czerwień rutenową.87 Istotnym krokiem w walidacji mechanotransdukcji przez kanały piezo jest zastosowanie metod in vivo w celu określenia funkcjonalnego znaczenia w sygnalizacji dotykowej. Poinformowano o Drosophila, gdzie usunięcie pojedynczego członka Piezo zmniejszyło reakcję mechaniczną na szkodliwe bodźce, nie wpływając na normalny dotyk.89 Chociaż ich struktura pozostaje do ustalenia, ta nowa rodzina białek mechanowrażliwych jest obiecującym tematem do przyszłych badań, poza granicami wrażenia dotykowego. Na przykład, niedawne badanie pacjentów z anemią (dziedziczną kserocytozą) pokazuje rolę Piezo 1 w utrzymywaniu homeostazy objętości erytrocytów.90
Transmembranowy podobny do kanału (TMC)
Niedawne badania wskazują, że dwa białka, TMC1 i TMC2, są niezbędne do mechanotransdukcji komórek włoskowatych.91 U ludzi i myszy odnotowano dziedziczną głuchotę spowodowaną mutacją genu TMC1 Obecność tych kanałów nie została jeszcze wykazana w układzie somatosensorycznym , ale wydaje się, że to dobry trop do zbadania.
Białko podobne do stomatyny 3 (SLP3)
Wykazano, że oprócz kanałów transdukcji, pewne dodatkowe białka połączone z kanałem odgrywają rolę we wrażliwości na dotyk. SLP3 ulega ekspresji w neuronach DRG ssaków. Badania na zmutowanych myszach pozbawionych SLP3 wykazały zmiany w prądach mechanosensacyjnych i mechanosensywnych.94,95 Dokładna funkcja SLP3 pozostaje nieznana. Może być łącznikiem między kanałem mechanowrażliwym a leżącymi pod nim mikrotubulami, jak zaproponowano dla jego homologu C. elegans MEC2.96 Ostatnio GR. Lewin lab zasugerował, że uwięź jest syntetyzowana przez neurony czuciowe DRG i łączy mechanowrażliwy kanał jonowy z macierzą pozakomórkową.97 Przerwanie połączenia znosi prąd mechanowrażliwy na RA, co sugeruje, że niektóre kanały jonowe są mechanowrażliwe tylko wtedy, gdy są uwiązane. Prądy mechanowrażliwe na RA są również hamowane przez lamininę-332, białko macierzy wytwarzane przez keratynocyty, co potwierdza hipotezę modulacji prądu mechanowrażliwego przez białka zewnątrzkomórkowe.98
Podrodzina kanałów K+
Równolegle do kationowych depolaryzujących mechanowrażliwych prądów badana jest obecność repolaryzujących mechanowrażliwych prądów K+. Kanały K+ w komórkach mechanowrażliwych mogą wpływać na równowagę prądu i przyczyniać się do określenia mechanicznego progu i przebiegu czasowego adaptacji mechanoreceptorów.
Członkowie KCNK należą do rodziny dwuporowych domen kanałów K+ (K2P).99,100 K2P wykazują niezwykły zakres regulacji przez czynniki komórkowe, fizyczne i farmakologiczne, w tym zmiany pH, ciepło, rozciąganie i deformację błony. Te K2P są aktywne w spoczynkowym potencjale błonowym. Kilka podjednostek KCNK ulega ekspresji w neuronach somatosensorycznych.101 Kanały KCNK2 (TREK-1), KCNK4 (TRAAK) i TREK-2 należą do nielicznych kanałów, dla których wykazano bezpośrednie bramkowanie mechaniczne przez rozciąganie błony.
Myszy z uszkodzonym genem KCNK2 wykazywały zwiększoną wrażliwość na ciepło i łagodne bodźce mechaniczne, ale normalny próg wycofania wobec szkodliwego nacisku mechanicznego wywieranego na tylną łapę przy użyciu testu Randalla Selitto.104 Myszy z niedoborem KCNK2 wykazują również zwiększoną przeczulicę termiczną i mechaniczną w stanach zapalnych warunki. Myszy z nokautem KCNK4 były nadwrażliwe na łagodną stymulację mechaniczną, a nadwrażliwość ta została zwiększona przez dodatkową inaktywację KCNK2.105. Zwiększona mechanowrażliwość tych myszy z nokautem może oznaczać, że rozciąganie normalnie aktywuje zarówno depolaryzujące, jak i repolaryzujące prądy mechanowrażliwe w skoordynowany sposób, podobnie jak brak równowagi depolaryzujące i repolaryzujące prądy bramkowane napięciem.
KCNK18 (TRESK) jest głównym czynnikiem wpływającym na przewodnictwo K+ w tle, które reguluje spoczynkowy potencjał błonowy neuronów somatosensorycznych.106 Chociaż nie wiadomo, czy KCNK18 jest bezpośrednio wrażliwy na stymulację mechaniczną, może odgrywać rolę w pośredniczeniu odpowiedzi na lekki dotyk, a także bolesne bodźce mechaniczne. Uważa się, że KCNK18 i w mniejszym stopniu KCNK3 są molekularnym celem hydroksy-a-sanshoolu, związku występującego w ziarnach pieprzu Schezuan, który aktywuje receptory dotykowe i wywołuje uczucie mrowienia u ludzi.107,108
Zależny od napięcia kanał K+ KCNQ4 (Kv7.4) ma kluczowe znaczenie dla ustawienia preferencji prędkości i częstotliwości subpopulacji szybko adaptujących się mechanoreceptorów zarówno u myszy, jak iu ludzi. Mutacja KCNQ4 była początkowo powiązana z postacią dziedzicznej głuchoty. Co ciekawe, niedawne badanie lokalizuje KCNQ4 w zakończeniach nerwów obwodowych skóry, szybko adaptujących się mieszków włosowych i ciałku Meissnera. W związku z tym utrata funkcji KCNQ4 prowadzi do selektywnego zwiększenia wrażliwości mechanoreceptora na drgania o niskiej częstotliwości. Warto zauważyć, że osoby z późnym ubytkiem słuchu z powodu dominujących mutacji genu KCNQ4 wykazują lepszą wydajność w wykrywaniu wibracji o małej amplitudzie i niskiej częstotliwości.109
Wgląd doktora Alexa Jimeneza
Dotyk jest uważany za jeden z najbardziej złożonych zmysłów w ludzkim ciele, zwłaszcza że nie ma za nim określonego organu. Zamiast tego zmysł dotyku odbywa się za pośrednictwem receptorów czuciowych, znanych jako mechanoreceptory, które znajdują się na skórze i reagują na nacisk mechaniczny lub zniekształcenie. Istnieją cztery główne typy mechanoreceptorów w skórze bezwłosej ssaków: ciałka blaszkowate, ciałka dotykowe, zakończenia nerwowe Merkla i ciałka bulwiaste. Mechanoreceptory działają w celu umożliwienia wykrywania dotyku, monitorowania położenia mięśni, kości i stawów, tzw. propriocepcji, a nawet wykrywania dźwięków i ruchu ciała. Zrozumienie mechanizmów budowy i funkcji tych mechanoreceptorów jest podstawowym elementem stosowania terapii i terapii w leczeniu bólu.
Wnioski
Dotyk jest zmysłem złożonym, ponieważ reprezentuje różne właściwości dotykowe, a mianowicie wibrację, kształt, fakturę, przyjemność i ból, z różnymi rozróżniającymi działaniami. Do tej pory związek między narządem dotykowym a zmysłem psychofizycznym był skorelowany, a specyficzne dla klasy markery molekularne dopiero się pojawiają. Obecnie potrzebne jest opracowanie testów na gryzoniach dopasowanych do różnorodności zachowań dotykowych, aby ułatwić przyszłą identyfikację genomiczną. Wykorzystanie myszy pozbawionych określonych podzbiorów czuciowych typów aferentnych znacznie ułatwi identyfikację mechanoreceptorów i czuciowych włókien aferentnych związanych z określoną modalnością dotykową. Co ciekawe, niedawna praca otwiera ważne pytanie o podłoże genetyczne cech mechanosensorycznych u człowieka i sugeruje, że mutacja pojedynczego genu może negatywnie wpływać na wrażliwość na dotyk.110 Podkreśla to, że patofizjologia ludzkiego deficytu dotyku jest w dużej mierze nieznana i z pewnością byłaby postęp poprzez precyzyjne zidentyfikowanie podzbioru neuronów czuciowych powiązanych z modalnością dotykową lub deficytem dotyku.
W zamian poczyniono postępy w określaniu właściwości biofizycznych prądów bramkowanych mechanicznie64. Rozwój w ostatnich latach nowych technik, pozwalających na monitorowanie zmian napięcia membrany, przy jednoczesnym rejestrowaniu prądów bramkowanych mechanicznie, okazał się cenną eksperymentalną metodą opisu prądy mechanoczułe o szybkiej, pośredniej i wolnej adaptacji (przegląd: Delmas i współpracownicy).66,111 W przyszłości będzie określenie roli obecnych właściwości w mechanizmach adaptacji funkcjonalnie zróżnicowanych mechanoreceptorów oraz wkładu mechanowrażliwych prądów K+ na pobudliwość LTMR i HTMR.
Molekularna natura prądów sterowanych mechanicznie u ssaków jest również obiecującym tematem badawczym w przyszłości. Przyszłe badania będą postępowały w dwóch perspektywach, po pierwsze, aby określić rolę cząsteczki pomocniczej, która wiąże kanały z cytoszkieletem i byłaby wymagana do nadania lub regulacji mechanowrażliwości kanałów jonowych, takich jak rodziny TRP i ASIC/EnaC. Po drugie, zbadanie dużego i obiecującego obszaru wkładu kanałów piezoelektrycznych poprzez udzielenie odpowiedzi na kluczowe pytania dotyczące mechanizmów przenikania i bramkowania, podzbioru neuronów czuciowych i modalności dotykowych z udziałem piezoelektrycznego oraz roli piezo w komórkach nieneuronalnych związanych z mechanosensacja.
Zmysł dotyku, w porównaniu do zmysłu wzroku, smaku, dźwięku i węchu, które wykorzystują określone narządy do przetwarzania tych wrażeń, może występować w całym ciele poprzez maleńkie receptory zwane mechanoreceptorami. Różne typy mechanoreceptorów można znaleźć w różnych warstwach skóry, gdzie mogą wykryć szeroki zakres stymulacji mechanicznej. W powyższym artykule opisano konkretne ciekawostki, które świadczą o postępie mechanizmów strukturalnych i funkcjonalnych mechanoreceptorów związanych ze zmysłem dotyku. Informacje podane w National Center for Biotechnology Information (NCBI). Zakres naszych informacji ogranicza się do chiropraktyki oraz urazów i stanów kręgosłupa. Aby omówić ten temat, prosimy zapytać dr Jimeneza lub skontaktować się z nami pod adresem�915-850-0900 .
Kurator: dr Alex Jimenez
Dodatkowe tematy: Back Pain
Na ból pleców jest jedną z najczęstszych przyczyn niepełnosprawności i nieobecności w pracy na całym świecie. W rzeczywistości, ból pleców został przypisany jako drugi najczęstszy powód wizyt u lekarza, przewyższający jedynie infekcje górnych dróg oddechowych. Około 80 procent populacji doświadczy pewnego rodzaju bólu pleców przynajmniej raz w ciągu całego życia. Kręgosłup jest złożoną strukturą zbudowaną z kości, stawów, więzadeł i mięśni oraz innych miękkich tkanek. Z tego powodu urazy i / lub pogorszenie warunków, takich jak przepukliny, może ostatecznie prowadzić do objawów bólu pleców. Urazy sportowe lub wypadki samochodowe są często najczęstszą przyczyną bólu pleców, jednak czasami najprostsze ruchy mogą mieć bolesne skutki. Na szczęście alternatywne opcje leczenia, takie jak opieka chiropraktyczna, mogą pomóc złagodzić ból pleców poprzez zastosowanie korekcji kręgosłupa i manualnych manipulacji, ostatecznie poprawiając ulgę w bólu.
1.�Moriwaki K, Yuge O. Cechy topograficzne skórnych nieprawidłowości w zakresie hipoestetyki i hiperestetyki skóry w przewlekłym bólu.Ból.�1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Cross Ref]
2.�Shim B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Uczulenie mechaniczne i cieplne nocyceptorów skóry u szczurów z doświadczalną neuropatią obwodowąNeuronauka.�2005;132: 193-201. doi: 10.1016/j.neuroscience.2004.12.036.�[PubMed][Cross Ref]
3.�Kleggetveit IP, J�rum E. Dysfunkcja dużych i małych włókien w urazach nerwów obwodowych z bólem samoistnym lub bez.J Ból.�2010;11:1305-10. doi: 10.1016/j.jpain.2010.03.004.�[PubMed][Cross Ref]
4.�Noback CR. Morfologia i filogeneza włosów.�Ann NY Acad Sci.�1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed][Cross Ref]
9.�Brown AG, Iggo A. Ilościowe badanie receptorów skórnych i włókien doprowadzających u kota i królika.J Physiol.�1967;193:707.�[Artykuł bezpłatny PMC][PubMed]
10.�Burgess PR, Petit D, Warren RM. Typy receptorów w owłosionej skórze kota dostarczane przez mielinowane włókna.�J Neurofizjol.�1968;31:833.�[PubMed]
11.�Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. Sox2-dodatnie komórki brodawki skórnej określają typ mieszka włosowego w naskórku ssaków.�Rozwój.�2009;136:2815. doi: 23/dev.10.1242.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
12.�mgr Husajna. Ogólny wzór ułożenia mieszków włosowych u szczura i myszy.�J Anat.�1971;109:307.�[Artykuł bezpłatny PMC][PubMed]
13.�Vielkind U, Hardy MH. Zmieniające się wzorce cząsteczek adhezji komórek podczas rozwoju mieszków włosowych sierści myszy. 2. Morfogeneza mieszków włosowych u mutantów włosowych, pręgowany i puszysty.�Acta Anat (Bazylea)�1996;157:183. doi: 94/10.1159.�[PubMed][Cross Ref]
14.�Hardy MH, Vielkind U. Zmieniające się wzorce cząsteczek adhezji komórek podczas rozwoju mieszków włosowych myszy. 1. Morfogeneza pęcherzyków u myszy typu dzikiego.�Acta Anat (Bazylea)�1996;157:169. doi: 82/10.1159.�[PubMed][Cross Ref]
15.�Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S i in. Funkcjonalna organizacja skórnych niskoprogowych neuronów mechanosensorycznychKomórka.�2011;147:1615. doi: 27/j.cell.10.1016.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
16.�Brown AG, Iggo A. Ilościowe badanie receptorów skórnych i włókien doprowadzających u kota i królika.J Physiol.�1967;193:707.�[Artykuł bezpłatny PMC][PubMed]
17.�Burgess PR, Petit D, Warren RM. Typy receptorów w owłosionej skórze kota dostarczane przez mielinowane włókna.�J Neurofizjol.�1968;31:833.�[PubMed]
18.�Vallbo A, Olausson H, Wessberg J, Norrsell U. System niezmielinizowanych aferentnych do nieszkodliwego mechanorecepcji w ludzkiej skórze.Mózg Res.�1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed][Cross Ref]
19.�Vallbo AB, Olausson H, Wessberg J. Niemielinizowane aferenty stanowią drugi system kodujący bodźce dotykowe ludzkiej skóry owłosionej.J Neurofizjol.�1999;81:2753.�[PubMed]
22.�Wessberg J, Olausson H, Fernström KW, Vallbo AB. Właściwości pola receptywnego niezmielinizowanych włókien aferentnych w ludzkiej skórzeJ Neurofizjol.�2003;89:1567. doi: 75/jn.10.1152.�[PubMed][Cross Ref]
23.�Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Molekularna wizualizacja genetyczna rzadkiego podzbioru niezmielinizowanych neuronów czuciowych, które mogą wykrywać delikatny dotyk.�Nat Neurosci.�2007;10:946. doi: 8/nn10.1038.�[PubMed][Cross Ref]
24.�Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G, et al. Niezmielinizowane włókna aferentne dotykowe sygnalizują dotyk i rzutują na korę wyspową.�Nat Neurosci.�2002;5:900. doi: 4/nn10.1038.[PubMed][Cross Ref]
25.�Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. Neurofizjologia niezmielinizowanych włókien aferentnych.Neurosci Biobehav Rev.�2010;34:185. doi: 91/j.neubiorev.10.1016.�[Recenzja][PubMed][Cross Ref]
26.�Kr�mer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M i in. Aktywacja korowej sieci bólowej poprzez miękką stymulację dotykową po wstrzyknięciu sumatryptanu.�Ból.�2007;133:72. doi: 8/j.pain.10.1016.�[PubMed][Cross Ref]
27.�Applebaum AE, Beall JE, Foreman RD, Willis WD. Pola organizacyjne i receptywne neuronów układu spinothalamicznego naczelnych.�J Neurofizjol.�1975;38:572.�[PubMed]
28.�Biały JC, słodki WH. Skuteczność chordotomii w bólu fantomowym po amputacji.�AMA Arch Neurol Psychiatria.�1952;67:315.�[PubMed]
29.�Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel i jego „Komórka Merkla”, morfologia, rozwój i fizjologia: przegląd i nowe wyniki.Anat Rec A Discov Mol Cell Evol Biol.�2003;271:225. doi: 39/ar.a.10.1002.�[PubMed][Cross Ref]
30.�Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Komórki Merkel ssaków pochodzą z linii naskórka.�Dev Biol.�2009;336:76. doi: 83/j.ydbio.10.1016.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
31.�Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N, et al. Komórki progenitorowe naskórka dają początek komórkom Merkla podczas rozwoju embrionalnego i homeostazy dorosłych.�J Komórka Biol.�2009;187:91. doi: 100/jcb.10.1083.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
32.�Ebara S, Kumamoto K, Baumann KI, Halata Z. Trójwymiarowe analizy kopuł dotykowych w owłosionej skórze kociej łapy ujawniają morfologiczne podłoża do złożonego przetwarzania sensorycznego.Neurosci Res.�2008;61:159. doi: 71/j.neures.10.1016.�[PubMed][Cross Ref]
33.�Guinard D, Usson Y, Guillermet C, Saxod R. Merkel kompleksy ludzkiej skóry cyfrowej: trójwymiarowe obrazowanie za pomocą konfokalnej mikroskopii laserowej i podwójnej immunofluorescencji.J Comp Neurol.�1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Cross Ref]
34.�Reinisch CM, Tschachler E. Kopuła dotykowa w ludzkiej skórze jest zasilana przez różne rodzaje włókien nerwowych.Anna Neurol.�2005;58:88. doi: 95/ana.10.1002.�[PubMed][Cross Ref]
35.�Maricich SM, Morrison KM, Mathes EL, Brewer BM. Gryzonie polegają na komórkach Merkel do zadań związanych z rozróżnianiem tekstur.�J. Neurosci.�2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
36.�Ikeda I, Yamashita Y, Ono T, Ogawa H. Selektywne fototoksyczne niszczenie szczurzych komórek Merkel znosi reakcje powoli adaptujących się jednostek mechanoreceptorów typu I.�J Physiol.�1994;479:247.�[Artykuł bezpłatny PMC][PubMed]
37.�Maricich SM, Wellnitz SA, Nelson AM, Leśniak DR, Gerling GJ, Lumpkin EA, et al. Komórki Merkla są niezbędne do lekkich reakcji na dotyk.�Nauka.�2009;324:1580. doi: 2/science.10.1126.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
38.�Diamond J, Holmes M, Pielęgniarka CA. Czy synapsy wzajemne komórek Merkla i neurytów są zaangażowane w inicjację reakcji dotykowych w skórze salamandry?J Physiol.�1986;376:101.�[Artykuł bezpłatny PMC][PubMed]
39.�Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Zależne od napięcia prądy w izolowanych pojedynczych komórkach Merkla szczurów.J Physiol.�1992;450:143.�[Artykuł bezpłatny PMC][PubMed]
40.�Wellnitz SA, Leśniak DR, Gerling GJ, Lumpkin EA. Regularność ciągłego wypalania ujawnia dwie populacje powoli adaptujących się receptorów dotykowych w mysiej owłosionej skórze.�J Neurofizjol.�2010;103:3378. doi: 88/jn.10.1152.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
41.�Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Immunolokalizacja akwaporyny-1 w mechanoreceptywnych zakończeniach Ruffini w więzadle przyzębia.Mózg Res.�2007;1157:32. doi: 40/j.brainres.10.1016.�[PubMed][Cross Ref]
42.�Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K, et al. Wykrywanie wyczuwającego kwas kanału jonowego 3 (ASIC3) w przyzębnych końcówkach Ruffini mysich siekaczy.�Neurosci Lett.�2011;488:173. doi: 7/j.neulet.10.1016.�[PubMed][Cross Ref]
43.�Johnsona KO. Role i funkcje mechanoreceptorów skóry.�Curr Opin Neurobiol.�2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed][Cross Ref]
44.�Wende H, Lechner SG, Cheret C, Bourane S, Kolańczyk ME, Pattyn A i in. Czynnik transkrypcyjny c-Maf kontroluje rozwój i funkcję receptora dotykowego.�Nauka.�2012;335:1373. doi: 6/science.10.1126.�[PubMed][Cross Ref]
45.�Mendelson M, Lowenstein WR. Mechanizmy adaptacji receptorów.�Nauka.�1964;144:554. doi: 5/science.10.1126.�[PubMed][Cross Ref]
46.�Loewenstein WR, Mendelson M. Składniki adaptacji receptora w ciałku Pacynina.�J Physiol.�1965;177:377.�[Artykuł bezpłatny PMC][PubMed]
47.�Pawson L, Prestia LT, Mahoney GK, G��l� B, Cox PJ, Pack AK. Interakcja GABAergiczna/glutaminergiczna-glejowa/neuronowa przyczynia się do szybkiej adaptacji w krwinkach pacyniańskich.J. Neurosci.�2009;29:2695�705. doi: 10.1523/JNEUROSCI.5974-08.2009.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
49.�Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A, et al. Niskoprogowe podtypy mechanoreceptorów selektywnie wyrażają MafA i są określone przez sygnalizację Ret.�Neuron.�2009;64:857. doi: 70/j.neuron.10.1016.�[PubMed][Cross Ref]
50.�Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. Rola sygnalizacji czynnika transkrypcyjnego Runx w różnicowaniu neuronów czuciowych zwoju korzenia grzbietowego.Neuron.�2006;49:379. doi: 93/j.neuron.10.1016.�[PubMed][Cross Ref]
51.�Luo W, Enomoto H, Ryż FL, Milbrandt J, Ginty DD. Molekularna identyfikacja szybko adaptujących się mechanoreceptorów i ich rozwojowa zależność od sygnalizacji ret.�Neuron.�2009;64:841. doi: 56/j.neuron.10.1016.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
52.�Vallbo AB, Hagbarth KE. Aktywność z mechanoreceptorów skóry rejestrowana przezskórnie u obudzonych ludziExp Neurol.�1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed][Cross Ref]
53.�Macefield VG. Charakterystyka fizjologiczna niskoprogowych mechanoreceptorów w stawach, mięśniach i skórze u ludziClin Exp Pharmacol Physiol.�2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed][Cross Ref]
54.�Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Fale Ca2+ w keratynocytach są przekazywane do neuronów czuciowych: zaangażowanie zewnątrzkomórkowej aktywacji receptora ATP i P2Y2.Biochem J.�2004;380:329. doi: 38/BJ10.1042.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
55.�Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. Sygnalizacja ATP ma kluczowe znaczenie dla odpowiedzi ludzkich keratynocytów na mechaniczną stymulację szokiem hipoosmotycznym.Exp Dermatol.�2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed][Cross Ref]
56.�Amano M, Fukata Y, Kaibuchi K. Regulacja i funkcje kinazy związanej z Rho.�Exp komórki Res.�2000;261:44. doi: 51/excr.10.1006.�[Przegląd]�[PubMed][Cross Ref]
57.�Koyama T, Oike M, Ito Y. Zaangażowanie kinazy Rho i kinazy tyrozynowej w uwalnianiu ATP wywołanym stresem hipotonicznym w komórkach śródbłonka aorty bydlęcej.J Physiol.�2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
59.�McCarter GC, Reichling DB, Levine JD. Transdukcja mechaniczna przez neurony zwoju korzenia grzbietowego szczura in vitroNeurosci Lett.�1999;273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[PubMed][Cross Ref]
60.�Drew LJ, Wood JN, Cesare P. Wyraźne właściwości mechanowrażliwe neuronów czuciowych wrażliwych i niewrażliwych na kapsaicynę.J. Neurosci.�2002;22:RC228.�[PubMed]
61.�Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P i in. Kanały jonowe wykrywające kwasy ASIC2 i ASIC3 nie przyczyniają się do mechanicznie aktywowanych prądów w neuronach czuciowych ssaków.J Physiol.�2004;556:691. doi: 710/jphysiol.10.1113.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
63.�Coste B, Crest M, Delmas P. Farmakologiczne rozwarstwienie i dystrybucja NaN/Nav1.9, prądów Ca2+ typu T i prądów kationów aktywowanych mechanicznie w różnych populacjach neuronów DRG.J Gen Physiol.�2007;129:57. doi: 77/jgp.10.1085.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
64.�Hao J, Delmas P. Wiele mechanizmów odczulania kanałów mechanoprzekaźnika kształtuje wypalanie neuronów mechanosensorycznych.J. Neurosci.�2010;30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[PubMed][Cross Ref]
65.�Drew LJ, Drewno JN. FM1-43 jest trwałym blokerem mechanowrażliwych kanałów jonowych w neuronach czuciowych i hamuje reakcje behawioralne na bodźce mechaniczne.Mol ból.�2007;3:1. doi: 10.1186/1744-8069-3-1.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
66.�Hao J, Delmas P. Rejestracja prądów mechanowrażliwych za pomocą mechanostymulatora napędzanego piezoelektrycznie.Protokół Nat.�2011;6: 979. doi: 90/nprot.10.1038.�[PubMed][Cross Ref]
75.�Strona AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ i in. Kanał jonowy ASIC1 przyczynia się do funkcji mechanoreceptora trzewnego, ale nie skórnego.�Gastroenterologia.�2004;127:1739. doi: 47/j.gastro.10.1053.�[PubMed][Cross Ref]
76.�Cena MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, et al. Kanał kationowy DRASIC przyczynia się do wykrywania dotyku skóry i bodźców kwasowych u myszy.�Neuron.�2001;32:1071. doi: 83/S10.1016-0896(6273)01-00547.�[Errata w: Neuron 5 Jul 2002;18] [35]�[PubMed][Cross Ref]
77.�Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M i in. Wycięcie kanału ASIC2 u myszy nie zaburza mechanosensacji skórnej, mechanonocycepcji trzewnej i słuchu.J Physiol.�2004;558:659. doi: 69/jphysiol.10.1113.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
78.�Damann N, Voets T, Nilius B. TRP w naszych zmysłach.�Curr Biol.�2008;18:R880. doi: 9/j.cub.10.1016.�[Przegląd]�[PubMed][Cross Ref]
79.�Christensen AP, Corey DP. Kanały TRP w mechanosensacji: aktywacja bezpośrednia czy pośrednia?�Nat Rev Neurosci.�2007;8:510-21. doi: 10.1038/nrn2149.�[Przegląd]�[PubMed][Cross Ref]
80.�Liedtke W, Tobin DM, Bargmann CI, Friedman JM. Ssak TRPV4 (VR-OAC) kieruje reakcjami behawioralnymi na bodźce osmotyczne i mechaniczne u Caenorhabditis elegans.Proc Natl Acad Sci USA A.�2003;100(Suppl 2):14531. doi: 6/pnas.10.1073.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
81.�Suzuki M, Mizuno A, Kodaira K, Imai M. Upośledzone odczucie ciśnienia u myszy pozbawionych TRPV4.�J Biol Chem.�2003;278:22664. doi: 8/jbc.M10.1074.�[PubMed][Cross Ref]
82.�Liedtke W, Choe Y, Mart�-Renom MA, Bell AM, Denis CS, Sali A, et al. Kanał aktywowany osmotycznie z receptorem waniloidowym (VR-OAC), kandydat na osmoreceptor kręgowców.Komórka.�2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
83.�Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Przejściowy potencjał receptora waniloidowego 4 jest niezbędny w indukowanym chemioterapią bólu neuropatycznego u szczurówJ. Neurosci.�2004;24:4444. doi: 52/JNEUROSCI.10.1523-0242.�[Errata w: J Neurosci. 04.2004 czerwiec;2004] [24][PubMed][Cross Ref]
84.�Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, et al. TRPA1 pośredniczy w działaniach zapalnych środowiskowych czynników drażniących i proalgetycznychKomórka.�2006;124:1269. doi: 82/j.cell.10.1016.�[PubMed][Cross Ref]
85.�Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ i in. TRPA1 przyczynia się do nocycepcji zimnej, mechanicznej i chemicznej, ale nie jest niezbędna do transdukcji komórek włoskowatych.�Neuron.�2006;50:277. doi: 89/j.neuron.10.1016.�[PubMed][Cross Ref]
86.�Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 i Piezo2 są podstawowymi składnikami odrębnych, aktywowanych mechanicznie kanałów kationowych.�Nauka.�2010;330:55. doi: 60/science.10.1126.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
87.�Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS i in. Białka piezo to tworzące pory podjednostki mechanicznie aktywowanych kanałów.�Natura.�2012;483:176. doi: 81/nature10.1038.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
88.�Bae C, Sachs F, Gottlieb PA. Mechanowrażliwy kanał jonowy Piezo1 jest hamowany przez peptyd GsMTx4.�Biochemia.�2011;50:6295. doi: 300/bi10.1021q.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
89.�Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. Rola Drosophila Piezo w mechanicznej nocycepcji.Natura.�2012;483:209-12. doi: 10.1038/nature10801.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
90.�Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. Mutacje w białku mechanotransdukcji PIEZO1 są związane z dziedziczną kserocytozą.�Krew.�2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
91.�Kawashima Y, G�l�oc GS, Kurima K, Labay V, Lelli A, Asai Y, et al. Mechanotransdukcja w komórkach rzęsatych ucha wewnętrznego myszy wymaga transbłonowych genów podobnych do kanałów.�J Clin Invest.�2011;121:4796. doi: 809/JCI10.1172.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
92.�Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouchäne J, et al. TMC1, ale nie TMC2, jest odpowiedzialny za autosomalne recesywne niesyndromiczne upośledzenie słuchu w rodzinach tunezyjskich.Audiol Neurootol.�2008;13:213. doi: 8/10.1159.�[PubMed][Cross Ref]
93.�Manji SS, Miller KA, Williams LH, Dahl HH. Identyfikacja trzech nowych szczepów myszy z ubytkiem słuchu z mutacjami w genie Tmc1.�Jestem J Pathol.�2012;180:1560. doi: 9/j.ajpath.10.1016.[PubMed][Cross Ref]
94.�Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, et al. Białko z domeny stomatyny niezbędne do odczuwania dotyku u myszy.�Natura.�2007;445:206-9. doi: 10.1038/nature05394.�[PubMed][Cross Ref]
95.�Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, et al. Mechanotransdukcja neuronów czuciowych i stomatyn.�J Neurofizjol.�2007;98:3802. doi: 8/jn.10.1152.[PubMed][Cross Ref]
96.�Huang M, Gu G, Ferguson EL, Chalfie M. Białko podobne do stomatyny niezbędne do mechanosensacji u C. elegans.Natura.�1995;378:292�5. doi: 10.1038/378292a0.�[PubMed][Cross Ref]
97.�Hu J, Chiang LY, Koch M, Lewin GR. Dowód na uwięź białkową zaangażowaną w dotyk somatyczny.�EMBO J.�2010;29:855. doi: 67/emboj.10.1038.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
98.�Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L, et al. Laminin-332 koordynuje mechanotransdukcję i bifurkację stożka wzrostu w neuronach czuciowych.�Nat Neurosci.�2011;14:993. doi: 1000/nn.10.1038.�[PubMed][Cross Ref]
99.�Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, et al. TWIK-1, wszechobecny ludzki, słabo do wewnątrz rektyfikujący kanał K+ o nowatorskiej strukturze.�EMBO J.�1996;15: 1004 11.[Artykuł bezpłatny PMC][PubMed]
100.�Lesage F. Farmakologia neuronalnych kanałów potasowych tła.�Neurofarmakologia.�2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed][Cross Ref]
101.�Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE i in. Analiza dystrybucji ludzkich dwuporowych kanałów potasowych w tkankach ośrodkowego układu nerwowego i obwodowychBrain Res Mol Brain Res.�2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Cross Ref]
102.�Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor� E. Stymulacja mechaniczna lub kwasowa, dwa interaktywne tryby aktywacji kanału potasowego TREK-1.�J Biol Chem.�1999;274:26691. doi: 6/jbc.10.1074.�[PubMed][Cross Ref]
103.�Maingret F, Fosset M, Lesage F, Lazdunski M, Honor� E. TRAAK jest ssaczym neuronowym kanałem K+ bramkowanym mechanicznie.�J Biol Chem.�1999;274:1381. doi: 7/jbc.10.1074.�[PubMed][Cross Ref]
104.�Alloui A, Zimmermann K, Mamet J, Duprat F, Nol J, Chemin J, et al. TREK-1, kanał K+ zaangażowany w polimodalną percepcję bólu.�EMBO J.�2006;25:2368. doi: 76/sj.emboj.10.1038.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
105.�Nol J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, et al. Aktywowane mechanicznie kanały K+ TRAAK i TREK-1 kontrolują zarówno percepcję ciepła, jak i zimna.�EMBO J.�2009;28:1308. doi: 18/emboj.10.1038.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
106.�Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. Kanały K+ o dwóch porach w domenie TRESK stanowią istotny składnik prądów potasowych w tle w neuronach zwoju korzeni grzbietowych myszy.J Physiol.�2007;585:867. doi: 79/jphysiol.10.1113.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
107.�Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR i in. Ostre środki z papryki syczuańskiej pobudzają neurony czuciowe poprzez hamowanie dwuporowych kanałów potasowych.�Nat Neurosci.�2008;11:772. doi: 9/nn.10.1038.�[Artykuł bezpłatny PMC][PubMed][Cross Ref]
109.�Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM i in. Kanały KCNQ4 K(+) dostrajają mechanoreceptory do normalnego odczuwania dotyku u myszy i człowieka.�Nat Neurosci.�2012;15:138. doi: 45/nn.10.1038.�[PubMed][Cross Ref]
110.�Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG, et al. Genetyczna podstawa cech mechanosensorycznych u ludzi.�PLoS Biol.�2012;10:e1001318. doi: 10.1371/journal.pbio.1001318.[Artykuł bezpłatny PMC][PubMed][Cross Ref]
111.�Delmas P, Hao J, Rodat-Despoix L. Molekularne mechanizmy mechanotransdukcji w neuronach czuciowych ssaków.Nat Rev Neurosci.�2011;12:139. doi: 53/nrn10.1038.�[PubMed][Cross Ref]
Ból w mechanorecepcji: Według CDC „ponad 50% dorosłych Amerykanów (125 milionów) miało układ mięśniowo-szkieletowy” zaburzenia bólowe w 2012 roku.�
W 40 r. Ponad 2012% dorosłych z zespołem bólu mięśniowo-szkieletowego stosowało uzupełniające podejście zdrowotne z jakiegokolwiek powodu. Było to znacznie więcej niż stosowanie wśród osób bez zespołu bólu mięśniowo-szkieletowego (24.1%). Stosowanie komplementarnych metod zdrowotnych z jakiegokolwiek powodu wśród osób z bólem szyi lub problemami karku było ponad dwukrotnie wyższe niż wśród osób bez tych problemów.
„Wśród osób dorosłych z zespołem bólu mięśniowo-szkieletowego, stosowanie jakiegokolwiek uzupełniającego podejścia zdrowotnego było najwyższe wśród osób z bólem szyi lub problemami (50.6%), a następnie u osób z innymi problemami mięśniowo-szkieletowymi (46.2%)”.
`` Stosowanie komplementarnych metod zdrowotnych z jakiegokolwiek powodu wśród osób z bólem lub problemami karku było ponad dwukrotnie wyższe niż wśród osób bez tych problemów.
Mechanoreceptory są receptorami czuciowymi reagującymi na ciśnienie mechaniczne lub zniekształcenia.
Należą do nich receptory skórne dla dotyku, receptory monitorujące długość i napięcie mięśni, receptory słuchowe i przedsionkowe i inne.
TEORIA BÓLU Z BRAMAMI
Bezbolesne wejście zamyka bramę do bolesnego wejścia.
Zapobiega to przemieszczaniu się odczuć bólowych na wyższe poziomy korowe
Aferenty o małej średnicy (ból) hamują hamowanie bólu
Aferenty o dużej średnicy (drgania) mają tendencję do pobudzania hamowania bólu.
Teoria ta twierdzi, że włókna nie-nocyceptywne mogą zakłócać sygnały z włókien bólu, a zatem hamować ból.
A o dużej średnicy? włókna są nienocyceptywne (nie przenoszą bodźców bólowych) i hamują efekty wystrzału A? i włókna C.
DORSAL COLUMN MEDIAL LEMNISCAL PATHWAY
WYKORZYSTANIE PERIPHERAL MECHANORECEPTORÓW DO ZMIANY BÓLU POSTRZEGANIE
Jak możemy pomóc?
REFLEKS KODU
Podekscytowany neuron dośrodkowy stymuluje pobudzające interneurony, które z kolei stymulują odruchowe neurony ruchowe dostarczające biceps, mięsień ramienia, który zgina (zgina) staw łokciowy. Skurcz bicepsa odciąga rękę od gorącego pieca.
Aferentny neuron stymuluje także interneurony hamujące, które z kolei hamują neurony odprowadzające, które dostarczają triceps, aby zapobiec ich skurczowi. Ten rodzaj połączenia neuronalnego obejmujący stymulację podaży nerwów do jednego mięśnia i równoczesne hamowanie nerwów do jego antagonistycznego mięśnia jest znany jako wzajemne hamowanie.
Aferentny neuron wciąż stymuluje inne interneurony, które przenoszą sygnał do rdzenia kręgowego do mózgu poprzez wznoszącą się ścieżkę. Dopiero gdy impuls dociera do obszaru czuciowego kory, osoba jest świadoma bólu, jego lokalizacji i rodzaju bodźca. Co więcej, kiedy impuls dociera do mózgu, informacja może być przechowywana jako pamięć i osoba może pomyśleć o tym, co się stało.
TERAPIA OPARTA NA RECEPTORZE
Korekty
Aktywacja mechanoreceptorów stawów poprzez zmiany chiropraktyczne może modulować i `` przyćmiać '' postrzeganie włókien o mniejszej średnicy przez mózg.
Powtarzanie aktywacji wspólnych mechanoreceptorów może tworzyć pozytywną plastyczność w drogach dotleniających.
Pozytywna plastyczność może zatrzymać ból
Wibracja
Stymulacja wibracyjna przy określonych częstotliwościach może zmienić postrzeganie bólu
Powtórzenie aktywacji dysków Merkel i ciałek Meissnera może stworzyć pozytywną plastyczność ścieżek aferentnych.
Ponownie, pozytywna plastyczność może wyłączyć ból
WIBRACJE
Ten typ urządzenia wykorzystuje wibracje sinusoidalne i oferuje ciągłą, wybieraną amplitudę w zakresie 0-5.2 mm w zależności od pozycji stopy i wybieraną częstotliwością w zakresie 5-30 Hz.
Szkolenie WBV wydaje się skuteczną, bezpieczną i odpowiednią interwencją dla pracowników siedzących z przewlekłym bólem krzyża.
Homotopowa stymulacja wibracyjno-dotykowa spowodowała 40% redukcję bólu spowodowanego ciepłem we wszystkich grupach badanych. Rozproszenie uwagi nie wydawało się wpływać na eksperymentalne oceny bólu
Stymulacja wibro-dotykowa skutecznie rekrutowała mechanizmy przeciwbólowe nie tylko w NC, ale także u pacjentów z przewlekłymi bólami mięśniowo-szkieletowymi, w tym FM.
LEKKI DOTYK
Łącznie 44 zdrowych ochotników doświadczyło bólu cieplnego i bodźców optymalnych dla TK (powolne szczotkowanie) i nieoptymalnych (szybkie szczotkowanie lub wibracja). Zastosowano trzy różne paradygmaty eksperymentalne: Jednoczesne stosowanie bólu wywołanego ciepłem i stymulacji dotykowej (powolne szczotkowanie lub wibracje); Powolne szczotkowanie, nakładane w różnym czasie i w różnych odstępach czasu, poprzedzające ból cieplny; Wolne i szybkie szczotkowanie poprzedzające ból spowodowany ciepłem
U ludzi, główne obszary mózgu otrzymujące informacje C-LTMR należą do układu somatosensorycznego i wpływają na przetwarzanie sieci mózgowych, takich jak kora tylnego odcinka bocznego tylnego lub przyśrodkowa kora przedczołowa. Intensywność dotyku ukierunkowanego na CT jest kodowana w pierwotnej i wtórnej korze somatosensorycznej (S1 kontralateralna, dwustronna S2), podczas gdy przyjemność jest zakodowana w korze przedniej obręczy. C-LTMR aktywują również regiony biorące udział w przetwarzaniu nagrody (skorupa i korze oczodołowo-czołowej) oraz w przetwarzaniu bodźców społecznych (tylny bruzdy czasowe górne).
WSZYSTKO PERIPHERAL MA CENTRALNE KONSEKWENCJE
CASE STUDY
47-letni mężczyzna doznał lewego CVA w październiku 2017.
Od czasu wypadku nie poruszył prawą stroną ciała.
Przedstawiony naszej klinice, ponieważ chce wrócić do niej
BADANIA NAUKOWE FIZYCZNE
Dyzartria
Zmieniona percepcja bólu
Trudność z prostą matematyką
Flakcid na RUE i RLE
BADANIA NAUKOWE FIZYCZNE
Pacjent nie miał ruchu, dopóki nie zaczęliśmy testować odczuć i odruchów:
ALLODYNIA:Odnosi się do ośrodkowego uczulenia na ból (zwiększona odpowiedź neuronów) po normalnie niebolesnej, często powtarzającej się stymulacji.
Allodynia może prowadzić do wywoływania reakcji bólowej na bodźce, co zwykle nie powoduje bólu.
Temperatura lub bodźce fizyczne mogą powodować allodynię, która może przypominać pieczenie i często pojawia się po uszkodzeniu miejsca.
Alldynia różni się od hiperalgezji, skrajnej, przesadnej reakcji na bodziec, który zwykle jest bolesny.
Dlaczego zlokalizowane uszkodzenie lub uraz spowodowany urazem prowadzi do przewlekłego, nieuleczalnego bólu u niektórych pacjentów? Co odpowiada za przełożenie urazu miejscowego z ostrym bólem na stan bólu przewlekłego? Dlaczego niektóre rodzaje bólu reagują na leki przeciwzapalne i/lub leki, podczas gdy inne formy bólu wymagają opiatów?
Ból jest skomplikowanym procesem obejmującym zarówno obwodowy układ nerwowy (PNS), jak i ośrodkowy układ nerwowy (OUN). Uszkodzenie tkanki wyzwala PNS, który przekazuje sygnały przez rdzeń kręgowy do mózgu, w którym następuje odczuwanie bólu. Co jednak powoduje, że intensywne doświadczanie bólu przeradza się w nieustanne zjawisko? Czy można coś zrobić, aby temu zapobiec? Dowody wskazują, że przewlekły ból wynika z kombinacji mechanizmów, takich jak neurologiczne „wspomnienia” wcześniejszego bólu.
Nocycepcja: najprostsza ścieżka
Ból ostry lub nocyceptywny charakteryzuje się jako regularne odczuwanie dyskomfortu, który pojawia się w odpowiedzi na bardzo podstawowe uszkodzenia lub urazy. Jest ochronny, ostrzega nas, abyśmy oddalili się od źródła zniewagi i zajęli się traumą. Mechanizmy wywołujące ból nocyceptywny obejmują transdukcję, która rozszerza zewnętrzną stymulację urazową do aktywności elektrycznej w wyspecjalizowanych nerwach nocyceptywnych pierwotnych doprowadzających. Następnie nerwy doprowadzające przenoszą informację czuciową z PNS do OUN.
W OUN dane dotyczące bólu są przekazywane przez pierwotne neurony czuciowe do centralnych komórek projekcyjnych. Po przekazaniu informacji do wszystkich obszarów mózgu, które są odpowiedzialne za naszą percepcję, następuje faktyczne doznanie zmysłowe. Ból nocyceptywny jest stosunkowo prostą reakcją na szczególnie prosty, ostry bodziec. Ale mechanicy odpowiedzialni za ból nocyceptywny nie potrafią zidentyfikować zjawisk, takich jak ból, który utrzymuje się pomimo usunięcia lub uzdrowienia stymulacji, jak w przypadku bólu fantomowego kończyn.
Ból i reakcja zapalna
W przypadku poważniejszych obrażeń, takich jak rany chirurgiczne, uszkodzenie tkanki może wywołać reakcję zapalną. Jednak inne stany, zwłaszcza zapalenie stawów, mogą również charakteryzować się utrzymującymi się stanami zapalnymi związanymi z silnymi objawami bólu. Mechanizmy tego typu bólu związanego z uszkodzeniem tkanki i reakcją zapalną różnią się od wczesnego ostrzegania bólu nocyceptywnego.
Obserwując nacięcie lub miejsce innego uszkodzenia lub urazu, w układzie nerwowym dochodzi do kaskady nadpobudliwości. To zjawisko „nakręcania się” ciała zaczyna się na skórze, gdzie jest wzmacniane wzdłuż nerwów obwodowych i kończy się reakcją nadwrażliwości wzdłuż rdzenia kręgowego (róg grzbietowy) i mózgu. Komórki zapalne otaczają wówczas obszary uszkodzenia tkanek, a także wytwarzają cytokiny i chemokiny, substancje, które mają pośredniczyć w procesie gojenia i regeneracji tkanek. Ale te czynniki mogą być również uważane za drażniące i dostosowują właściwości pierwotnych neuronów czuciowych otaczających obszar urazu.
Zatem główne czynniki, które wywołują ból zapalny, obejmują uszkodzenie wysokoprogowych nocyceptorów, znanych jako sensytyzacja obwodowa, zmiany i zmiany neuronów w układzie nerwowym oraz wzmocnienie pobudliwości neuronów w OUN. Reprezentuje to uczulenie ośrodkowe i jest odpowiedzialne za nadwrażliwość, w której obszary sąsiadujące z obszarami prawdziwego urazu będą odczuwać ból tak, jakby zostały zranione. Tkanki te mogą również reagować na stymulację, która normalnie nie powoduje bólu, taką jak dotyk, noszenie ubrania, lekki ucisk, a nawet czesanie własnych włosów, tak jakby były naprawdę bolesne, określane jako allodynia.
Ból neuropatyczny wynika z uszkodzenia lub urazu układu nerwowego, takiego jak zespół cieśni nadgarstka, neuralgia popółpaścowa i neuropatia cukrzycowa. Chociaż niektóre mechanizmy, które wydają się powodować ból neuropatyczny, pokrywają się z mechanizmami odpowiedzialnymi za ból zapalny, wiele z nich jest innych, a zatem wymagają innego podejścia do ich leczenia.
Proces uczulenia obwodowego i ośrodkowego jest utrzymywany, przynajmniej teoretycznie i doświadczalnie, podczas pobudzającego neuroprzekaźnika, glutaminianu, który, jak się uważa, jest uwalniany, gdy aktywowany jest receptor N-metylo-D-asparaginianu (NMDA).
Układ nerwowy składa się z neuroprzekaźników hamujących lub pobudzających. Większość tego, co pozwala naszemu układowi nerwowemu odpowiednio reagować na uszkodzenia lub urazy, to dostrajanie lub hamowanie różnych procesów. Nadmierne pobudzenie układu nerwowego jest postrzegane jako problem w wielu różnych zaburzeniach. Na przykład nadmierna aktywacja receptora NMDA może być również związana z zaburzeniami afektywnymi, nieprawidłowościami współczulnym, a nawet tolerancją na opiaty.
Nawet zwykły ból nocyceptywny w pewnym stopniu aktywuje receptor NMDA i uważa się, że prowadzi do uwalniania glutaminianu. Niemniej jednak w bólu neuropatycznym kluczowa jest nadwrażliwość na receptor NMDA.
W przypadku innych rodzajów bólu przewlekłego, takich jak fibromialgia i napięciowe bóle głowy, niektóre mechanizmy działające w bólu zapalnym i neuropatycznym mogą również powodować podobne nieprawidłowości w układzie bólowym, w tym uczulenie ośrodkowe, wyższą pobudliwość szlaków somatosensorycznych i zmniejszenie mechanizmy hamowania ośrodkowego układu nerwowego.
Uczulenie obwodowe
Cyklooksygenaza (COX) również odgrywa istotną funkcję zarówno w uczuleniach obwodowych, jak i ośrodkowych. COX-2 jest jednym z enzymów indukowanych podczas procesu zapalnego; COX-2 przekształca kwas arachidonowy w prostaglandyny, które zwiększają czułość obwodowych zakończeń nocyceptorów. W rzeczywistości zapalenie obwodowe powoduje również wytwarzanie COX-2 z OUN. Sygnały z obwodowych nocyceptorów są częściowo odpowiedzialne za tę regulację w górę, ale wydaje się, że istnieje również element humoralny w transdukcji sygnałów bólu przez barierę krew-mózg.
Na przykład w modelach eksperymentalnych COX-2 jest generowana z OUN, nawet jeśli zwierzęta otrzymują blokadę nerwów czuciowych przed obwodową stymulacją zapalną. COX-2, który ulega ekspresji przez neurony rogu grzbietowego rdzenia kręgowego, uwalnia prostaglandyny, które działają na centralne lub presynaptyczne zakończenia nocyceptywnych włókien czuciowych w celu zwiększenia uwalniania przekaźnika. Dodatkowo działają postsynaptycznie na neurony rogu grzbietowego, powodując bezpośrednią depolaryzację. I wreszcie hamują aktywność receptora glicyny, a to jest przekaźnik hamujący. Dlatego prostaglandyny powodują wzrost pobudliwości neuronów centralnych.
Plastyczność mózgu i ośrodkowe uczulenie
Uczulenie ośrodkowe opisuje zmiany zachodzące w mózgu w reakcji na powtarzaną stymulację nerwów. Po powtarzających się bodźcach ilość hormonów i sygnałów elektrycznych w mózgu zmienia się, gdy neurony rozwijają „pamięć” do reagowania na te sygnały. Ciągła stymulacja tworzy potężniejszą pamięć mózgu, dzięki czemu mózg będzie reagował szybciej i skuteczniej, gdy będzie poddawany identycznej stymulacji w przyszłości. Wynikające z tego modyfikacje okablowania i reakcji mózgu określa się mianem plastyczności neuronalnej, która opisuje zdolność mózgu do łatwej zmiany samego siebie lub ośrodkowego sensytyzacji. Dlatego mózg jest aktywowany lub uczulany przez poprzednie lub powtarzane bodźce, aby stać się bardziej pobudliwym.
Wahania ośrodkowego sensytyzacji pojawiają się po wielokrotnych spotkaniach z bólem. Badania na zwierzętach wskazują, że wielokrotna ekspozycja na bolesną stymulację zmieni próg bólu zwierzęcia i doprowadzi do silniejszej reakcji na ból. Naukowcy uważają, że te modyfikacje mogą wyjaśniać uporczywy ból, który może wystąpić nawet po udanej operacji kręgosłupa. Chociaż przepuklina dysku może zostać usunięta z zaciśniętego nerwu, ból może nadal być wspomnieniem ucisku nerwu. Noworodki poddawane obrzezaniu bez znieczulenia będą silniej reagować na przyszłą bolesną stymulację, taką jak rutynowe zastrzyki, szczepienia i inne bolesne procesy. Te dzieci mają nie tylko wyższą reakcję hemodynamiczną, znaną jako tachykardia i tachypnea, ale również rozwiną się u nich wzmożony płacz.
Ta neurologiczna pamięć bólu była intensywnie badana. W raporcie na temat swoich poprzednich badań Woolf zauważył, że poprawiona pobudliwość odruchowa po uszkodzeniu lub urazie tkanek obwodowych nie zależy od ciągłych sygnałów wejściowych z obwodu; raczej, kilka godzin po urazie obwodowym, pola receptorowe rogów grzbietowych rdzenia kręgowego nadal się powiększały. Badacze udokumentowali również znaczenie rdzeniowego receptora NMDA w indukcji i utrzymywaniu sensytyzacji ośrodkowej.
Znaczenie dla zarządzania bólem
Po ustaleniu sensytyzacji ośrodkowej często wymagane są większe dawki leków przeciwbólowych, aby je stłumić. Analgezja zapobiegawcza lub terapia przed nasileniem się bólu może zmniejszyć wpływ wszystkich tych stymulacji na OUN. Woolf wykazał, że dawka morfiny wymagana do zatrzymania nadpobudliwości ośrodkowej, podana przed krótką, szkodliwą stymulacją elektryczną u szczurów, stanowiła jedną dziesiątą dawki wymaganej do zniesienia aktywności po jej wzroście. Przekłada się to na praktykę kliniczną.
W badaniu klinicznym z udziałem 60 pacjentek poddawanych histerektomii brzusznej osoby, które otrzymały 10 mg morfiny dożylnie w czasie indukcji znieczulenia, wymagały znacznie mniej morfiny w celu opanowania bólu pooperacyjnego. Ponadto wrażliwość na ból wokół rany, określana jako wtórna przeczulica bólowa, była również zmniejszona w grupie leczonej wstępnie morfiną. Analgezja zapobiegawcza była stosowana z porównywalnym sukcesem w różnych ustawieniach chirurgicznych, w tym operacji przednakrętowej i po ortopedycznej.
Pojedyncza dawka 40 lub 60 mg/kg paracetamolu doodbytniczego ma wyraźne działanie oszczędzające morfinę w chirurgii jednego dnia u dzieci, jeśli jest podawana w indukcji znieczulenia. Ponadto dzieci z wystarczającą analgezją za pomocą paracetamolu odczuwały znacznie mniej pooperacyjnych nudności i wymiotów.
Antagoniści receptora NMDA, podani przed operacją, wywierają działanie przeciwbólowe po operacji. W literaturze istnieją różne doniesienia popierające stosowanie ketaminy i dekstrometorfanu w okresie przedoperacyjnym. U pacjentów poddawanych rekonstrukcji więzadła krzyżowego przedniego, 24-godzinne kontrolowane przez pacjenta spożycie opioidów przeciwbólowych było znacznie mniejsze w przedoperacyjnej kategorii dekstrometorfanu w porównaniu z grupą placebo.
W badaniach z podwójnie ślepą próbą, kontrolowanych placebo, gabapentyna została wskazana jako lek przeciwbólowy w premedykacji u pacjentek poddawanych mastektomii i histerektomii. Przedoperacyjna doustna gabapentyna zmniejszyła ból i pooperacyjne spożycie środków przeciwbólowych bez luki w skutkach ubocznych w porównaniu z placebo.
Przedoperacyjne podanie niesteroidowych leków przeciwzapalnych (NLPZ) wykazało istotne zmniejszenie pooperacyjnego stosowania opioidów. COX-2 są preferowane ze względu na ich względny brak działania płytek krwi i istotny profil bezpieczeństwa żołądkowo-jelitowego w porównaniu z konwencjonalnymi NLPZ. Celekoksyb, rofekoksyb, waldekoksyb i parekoksyb, podawane poza Stanami Zjednoczonymi przed operacją, zmniejszają pooperacyjne zażywanie narkotyków o ponad 40 procent, przy czym wielu pacjentów używa mniej niż połowę opioidów w porównaniu z placebo.
Wydaje się, że blokowanie przewodnictwa nerwowego w okresie przedoperacyjnym zapobiega rozwojowi sensytyzacji ośrodkowej. Zespół kończyny fantomowej (PLS) przypisano zjawisku zwichnięcia kręgosłupa. Pacjenci z amputacją
często mają usunięte pieczenie lub mrowienie w części ciała. Jedną z możliwych przyczyn jest to, że włókna nerwowe w kikucie są stymulowane, a mózg interpretuje sygnały jako pochodzące z amputowanej części. Drugim jest przegrupowanie w obszarach korowych, tak że obszar, powiedzmy, dla ręki, teraz reaguje na sygnały z innych części ciała, ale nadal interpretuje je jako nadchodzące po amputowaną rękę.
Jednak w przypadku pacjentów poddawanych amputacji kończyny dolnej w znieczuleniu zewnątrzoponowym, żaden z 11 pacjentów, którzy otrzymali blokadę zewnątrzoponową lędźwiową z użyciem bupiwakainy i morfiny przez 72 godziny przed operacją, nie rozwinął PLS. W przypadku osób, które przeszły znieczulenie ogólne bez wcześniejszej blokady zewnątrzoponowej lędźwiowej, 5 z 14 pacjentów miało PLS po 6 tygodniach, a 3 nadal doświadczało PLS po 1 roku.
Woolf i Chong zauważyli, że doskonałe leczenie przedoperacyjne, śródoperacyjne i pooperacyjne obejmuje „NLPZ w celu zmniejszenia aktywacji/centralizacji nocyceptorów, miejscowe środki znieczulające blokujące napływ czuciowy oraz leki działające ośrodkowo, takie jak opiaty”. Zmniejszenie bólu okołooperacyjnego technikami zapobiegawczymi zwiększa satysfakcję, przyspiesza wydzielinę, oszczędza stosowanie opioidów, zmniejsza zaparcia, sedację, nudności i zatrzymanie moczu, a nawet może powstrzymać rozwój bólu przewlekłego. Anestezjolodzy i chirurdzy powinni rozważyć włączenie tych technik do swojej codziennej praktyki.
Gdy ból pojawia się w wyniku uszkodzenia lub urazu w wyniku operacji, rdzeń kręgowy może osiągnąć stan nadpobudliwości, w którym występują nadmierne reakcje bólowe, które mogą utrzymywać się przez kilka dni, tygodni, a nawet lat.
Dlaczego zlokalizowane uszkodzenie powstałe w wyniku urazu powoduje u niektórych pacjentów przewlekły, nieuleczalny ból? Uszkodzenie tkanki prowadzi do konstelacji zmian w pobudliwości kręgosłupa, w tym zwiększonego spontanicznego wyzwalania, większej amplitudy i długości odpowiedzi, zmniejszonego progu, zwiększonego wyładowania do powtarzanej stymulacji i rozszerzonego pola recepcyjnego. Utrzymywanie się tych zmian, które są zbiorczo określane jako sensytyzacja ośrodkowa, wydaje się mieć fundamentalne znaczenie dla przedłużonego zwiększania wrażliwości na ból, która definiuje ból przewlekły. Liczne leki i/lub leki, jak również miejscowa blokada nerwowa znieczulenia mogą ograniczać nasilenie nawrotu ośrodkowego układu nerwowego (OUN), o czym świadczy zmniejszenie bólu i zmniejszone zużycie opioidów w modelach z zapobiegawczymi lekami przeciwbólowymi.
Wgląd doktora Alexa Jimeneza
Opieka chiropraktyczna jest alternatywną opcją leczenia, która wykorzystuje korekty kręgosłupa i manipulacje manualne w celu bezpiecznego i skutecznego przywrócenia oraz utrzymania prawidłowego ustawienia kręgosłupa. Badania naukowe wykazały, że niewspółosiowość kręgosłupa lub podwichnięcia mogą prowadzić do przewlekłego bólu. Opieka chiropraktyczna jest powszechnie stosowana w leczeniu bólu, nawet jeśli objawy nie są związane z urazem i/lub stanem układu mięśniowo-szkieletowego i nerwowego. Starannie wyrównując kręgosłup, a kręgarz może pomóc zmniejszyć stres i nacisk ze struktur otaczających główny składnik podstawy naszego ciała, ostatecznie zapewniając ulgę w bólu.
Czynność i ból jelitowego układu nerwowego
Jeśli chodzi o zmniejszone stosowanie leków i/lub leków, w tym opioidów, w celu zapobiegania skutkom ubocznym, takim jak problemy ze zdrowiem przewodu pokarmowego, może odgrywać rolę właściwa funkcja jelitowego układu nerwowego.
Jelitowy układ nerwowy (ENS) lub wewnętrzny układ nerwowy jest jedną z kluczowych gałęzi autonomicznego układu nerwowego (ANS) i składa się z przypominającego siatkę układu nerwów, który moduluje rolę przewodu pokarmowego. Jest w stanie działać niezależnie od współczulnego i przywspółczulnego układu nerwowego, nawet jeśli może na niego wpływać. ENS można również nazwać drugim mózgiem. Pochodzi z komórek grzebienia nerwowego.
Jelitowy układ nerwowy u ludzi składa się z około 500 milionów neuronów, w tym licznych typów komórek Dogla, około jednej dwusetnej liczby neuronów w mózgu. Jelitowy układ nerwowy jest wprowadzany do wyściółki układu pokarmowego, zaczynając od przełyku i rozciągając się do odbytu. Komórki Dogla, znane również jako komórki Dogla, odnoszą się do pewnego rodzaju wielobiegunowych tkanek nadnerczy w zwojach współczulnych przedkręgowych.
ENS jest zdolny do autonomicznych funkcji, takich jak koordynacja odruchów; mimo że otrzymuje znaczne unerwienie w autonomicznym układzie nerwowym, działa i może działać niezależnie od mózgu i rdzenia kręgowego. Jelitowy układ nerwowy został opisany jako „drugi mózg” z wielu powodów. Jelitowy układ nerwowy może działać autonomicznie. Zwykle komunikuje się z ośrodkowym układem nerwowym (OUN) za pośrednictwem układu przywspółczulnego lub nerwu błędnego oraz układu współczulnego, czyli zwojów przedkręgowych, czyli układu nerwowego. Jednak badania na kręgowcach ujawniają, że po przecięciu nerwu błędnego jelitowy układ nerwowy nadal funkcjonuje.
U kręgowców jelitowy układ nerwowy obejmuje neurony odprowadzające, neurony doprowadzające i interneurony, z których wszystkie sprawiają, że jelitowy układ nerwowy jest w stanie przenosić odruchy i działać jako centrum integracyjne przy braku dopływu do ośrodkowego układu nerwowego. Neurony czuciowe informują o warunkach mechanicznych i chemicznych. Jelitowy układ nerwowy ma zdolność do zmiany odpowiedzi w oparciu o takie czynniki, jak skład składników odżywczych i objętościowych. Ponadto ENS zawiera komórki podporowe, które są bardzo podobne do astrogleju mózgu i barierę dyfuzyjną wokół naczyń włosowatych otaczających zwoje, która jest jak bariera krew-mózg naczyń krwionośnych.
Jelitowy układ nerwowy (ENS) odgrywa kluczową rolę w procesach zapalnych i nocyceptywnych. Leki i/lub leki wchodzące w interakcję z układem nerwowym jelit wzbudziły ostatnio duże zainteresowanie ze względu na ich zdolność do regulowania wielu aspektów fizjologii i patofizjologii jelit. W szczególności eksperymenty na zwierzętach wykazały, że receptory aktywowane przez proteinazy (PAR) mogą być niezbędne do neurogennego zapalenia w jelicie. Ponadto agoniści PAR2 wydają się wywoływać nadwrażliwość jelitową i stany hiperalgezy, co sugeruje rolę tego receptora w odczuwaniu bólu trzewnego.
Ponadto PAR, wraz z aktywującymi je proteinazami, stanowią nowe, ekscytujące cele interwencji terapeutycznej w układzie nerwowym jelit. Zakres naszych informacji ogranicza się do chiropraktyki oraz urazów i stanów kręgosłupa. W celu omówienia tematu, prosimy o kontakt z dr Jimenezem lub o kontakt pod adresem 915-850-0900 .
Kurator: dr Alex Jimenez
Dodatkowe tematy: rwa kulszowa
Na rwę kulszową jest medycznie określany jako zbiór objawów, a nie pojedynczy uraz i/lub stan. Objawy bólu nerwu kulszowego lub rwy kulszowej mogą różnić się częstotliwością i intensywnością, jednak najczęściej opisuje się je jako nagły, ostry (podobny do noża) lub elektryczny ból, który promieniuje z dolnej części pleców w dół pośladków, bioder, ud i nogi w stopę. Inne objawy rwy kulszowej mogą obejmować uczucie mrowienia lub pieczenia, drętwienie i osłabienie wzdłuż nerwu kulszowego. Rwa kulszowa najczęściej dotyka osoby w wieku od 30 do 50 lat. Często może rozwijać się w wyniku zwyrodnienia kręgosłupa związanego z wiekiem, jednak ucisk i podrażnienie nerwu kulszowego spowodowane wybrzuszeniem lub przepuklina dysku, oprócz innych problemów zdrowotnych kręgosłupa, może również powodować ból nerwu kulszowego.
Narzędzie Find A Practitioner firmy IFM to największa sieć skierowań w medycynie funkcjonalnej, stworzona, aby pomóc pacjentom zlokalizować lekarzy medycyny funkcjonalnej na całym świecie. Certyfikowani praktycy IFM są wymienieni na pierwszym miejscu w wynikach wyszukiwania ze względu na ich rozległe wykształcenie w zakresie medycyny funkcjonalnej

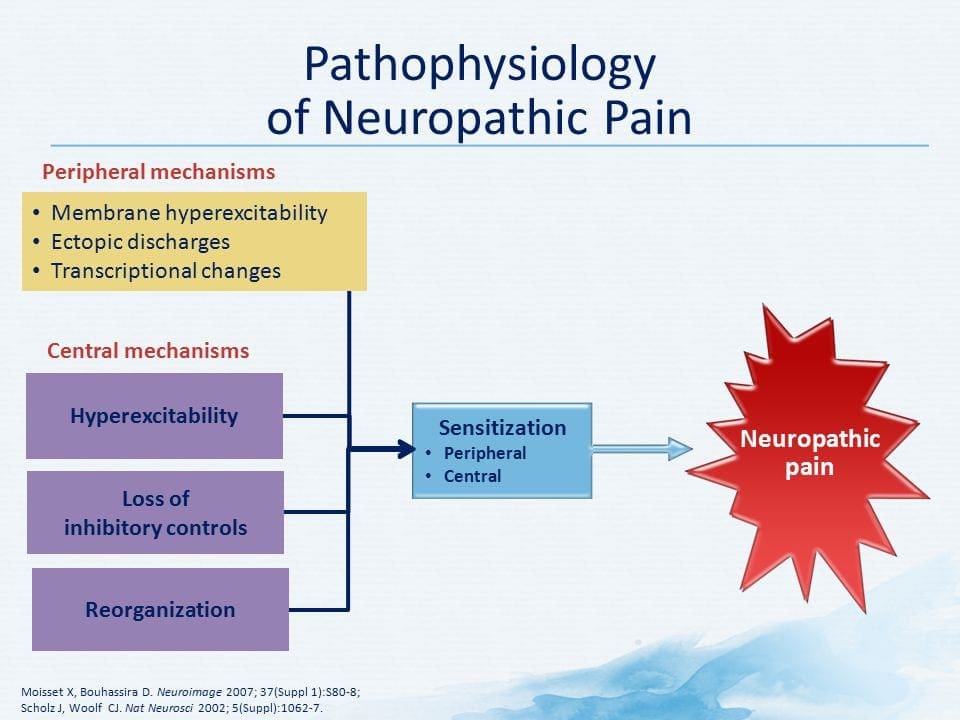
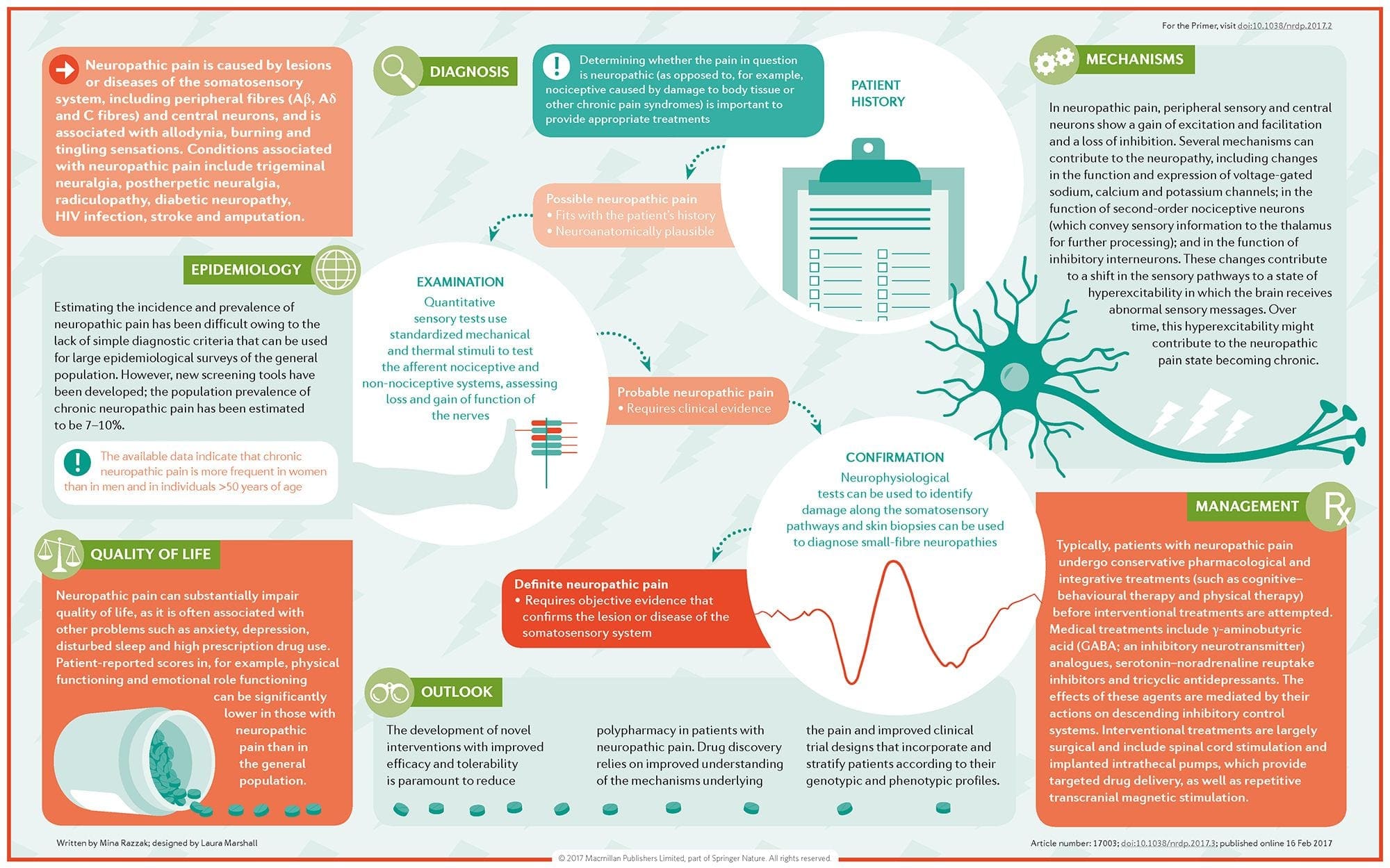




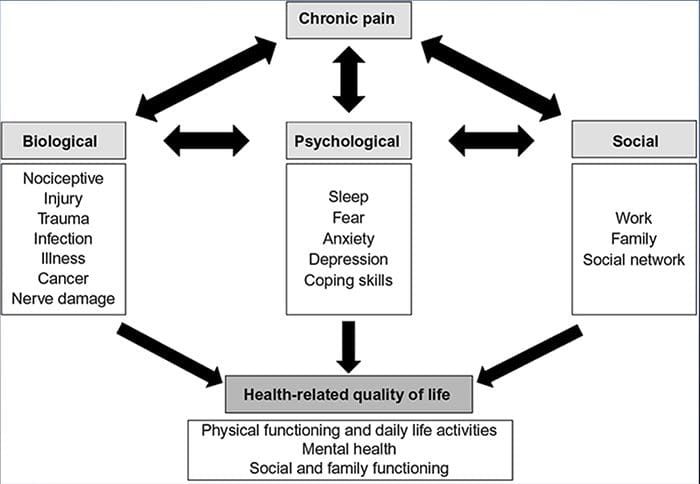
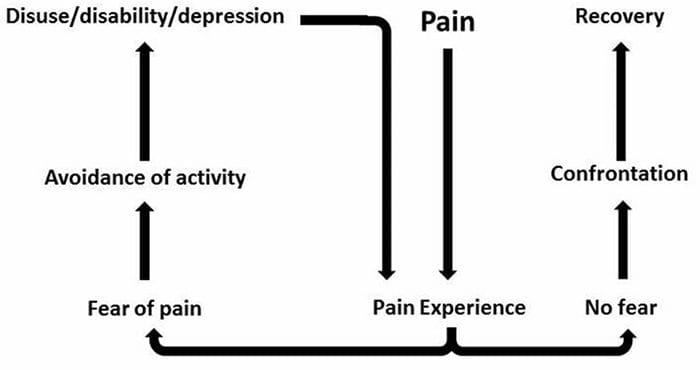
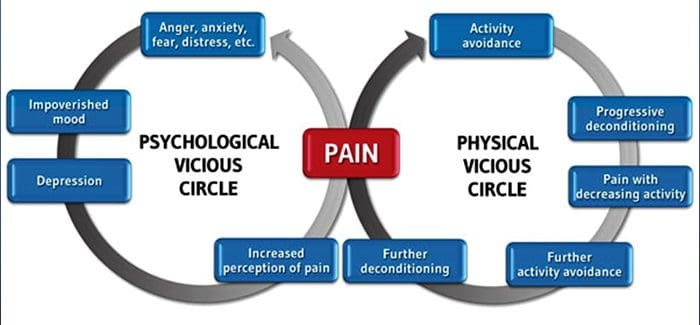


















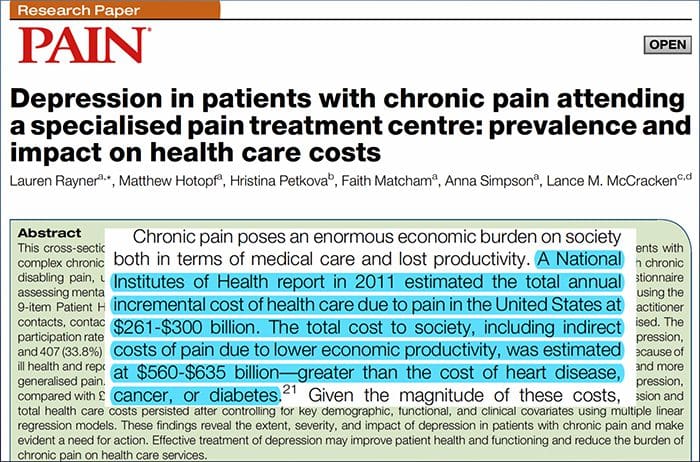









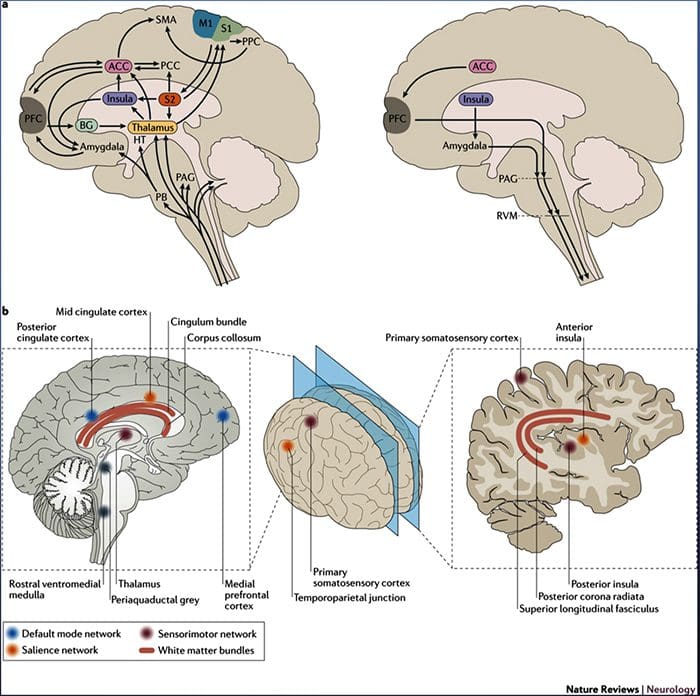


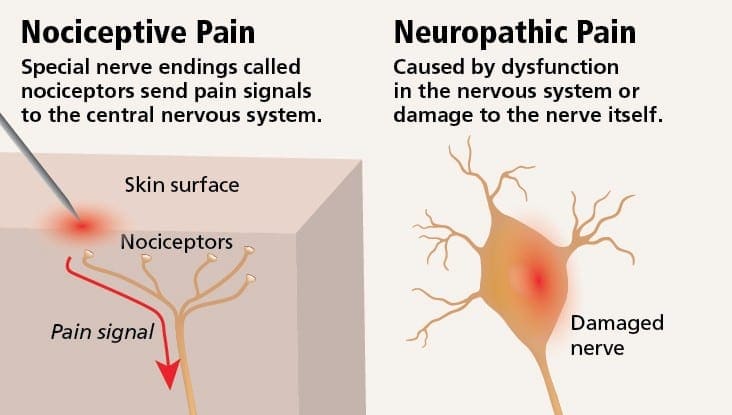
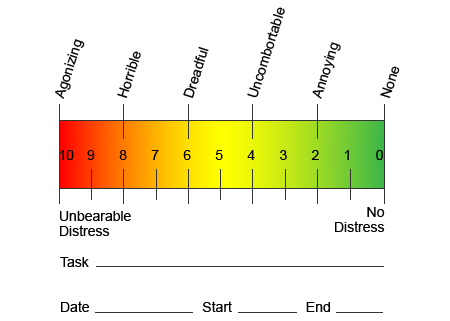


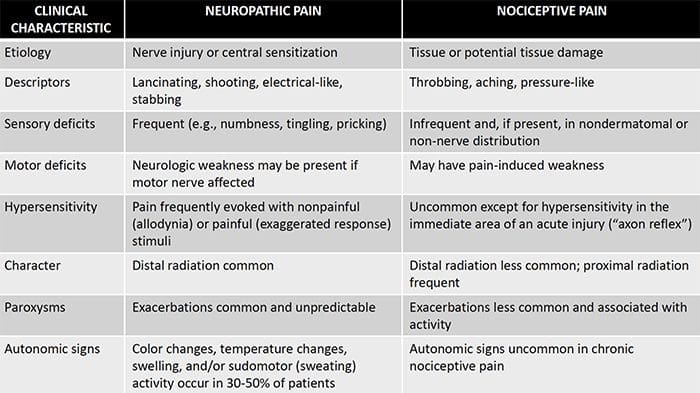
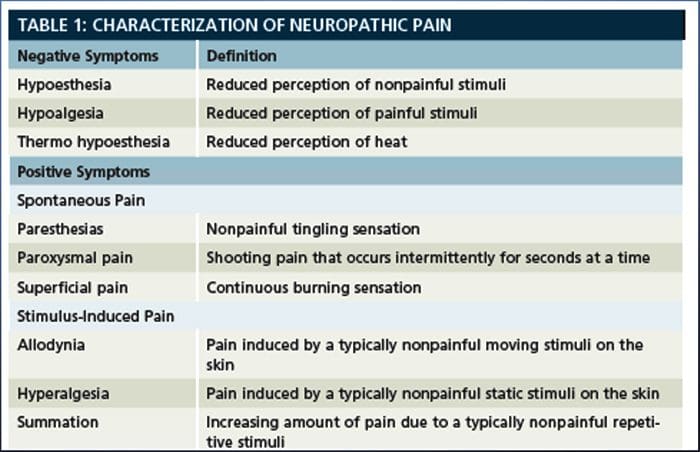
 PATOGENEZA BÓLU NEUROPATYCZNEGO
PATOGENEZA BÓLU NEUROPATYCZNEGO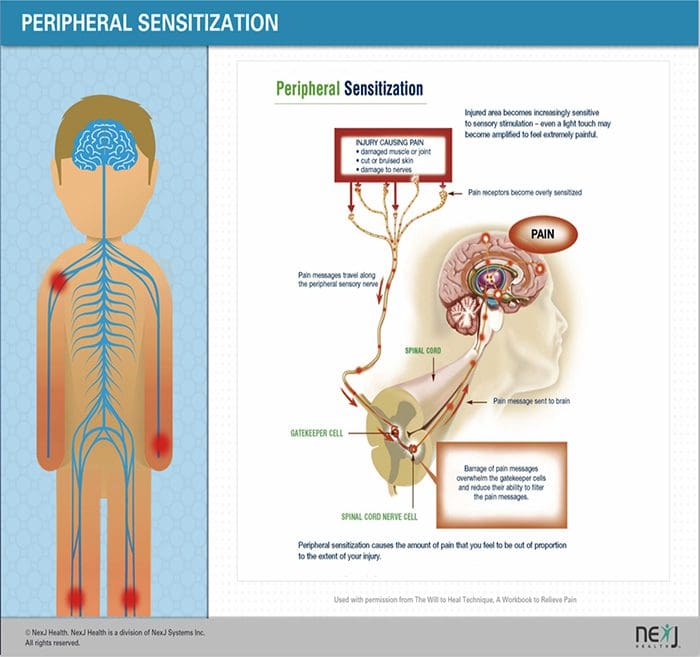
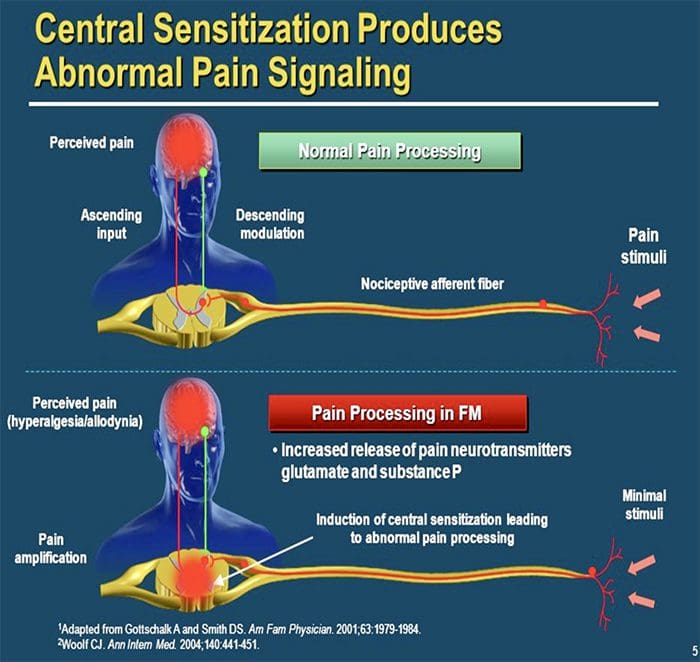
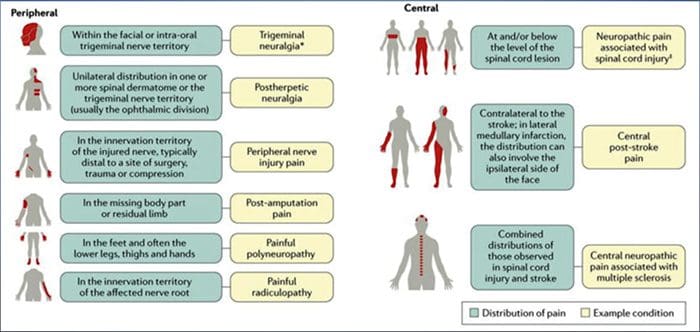 NAJCZĘSTSZE PRZYCZYNY
NAJCZĘSTSZE PRZYCZYNY
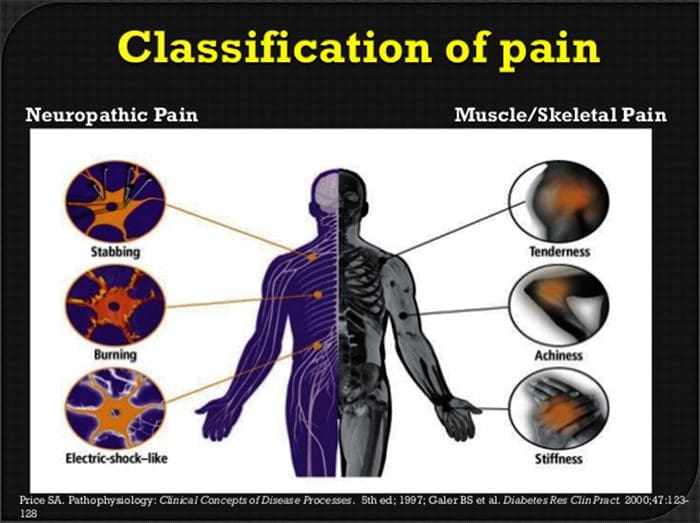
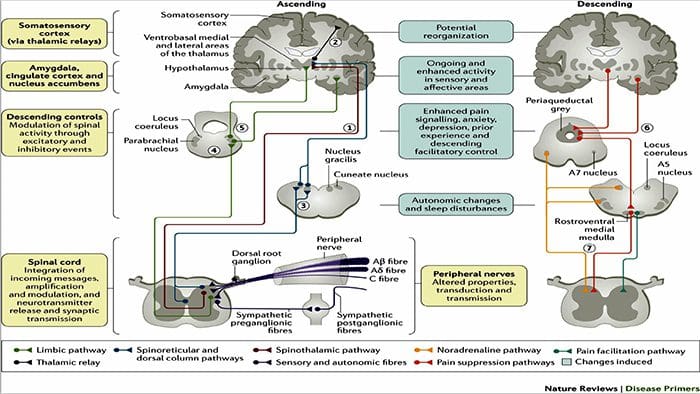
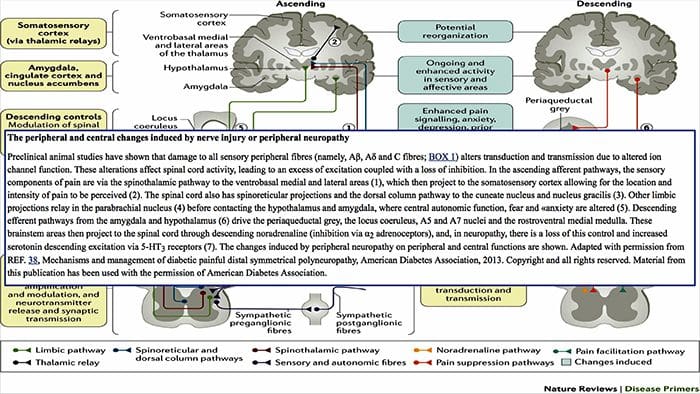

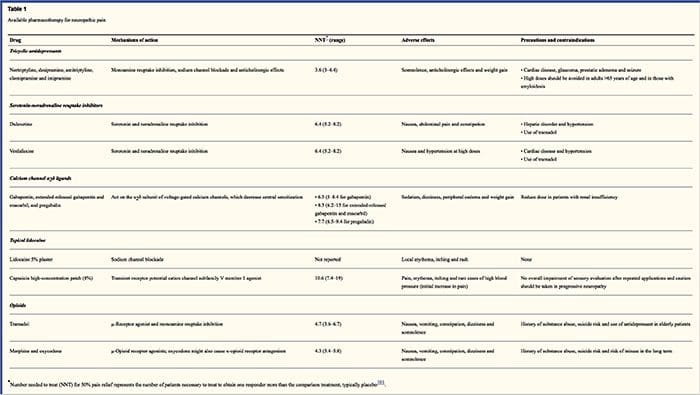



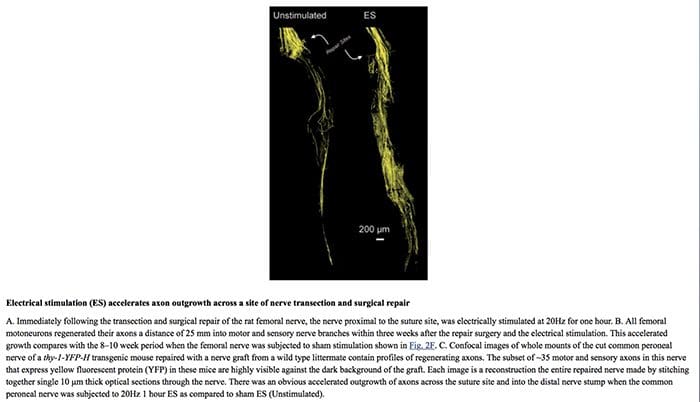

 BÓL PHANTOMOWY I RZECZYWISTOŚĆ ROZSZERZONA
BÓL PHANTOMOWY I RZECZYWISTOŚĆ ROZSZERZONA
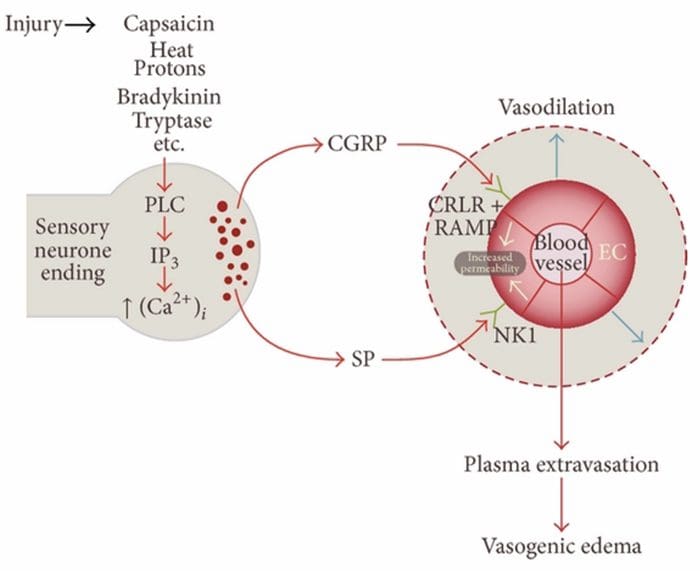 NAJCZĘSTSZE PRZYCZYNY
NAJCZĘSTSZE PRZYCZYNY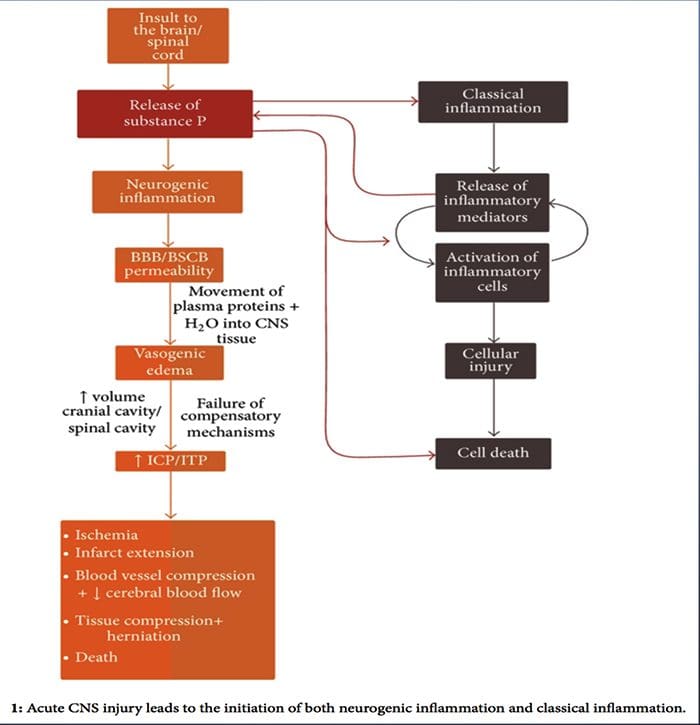


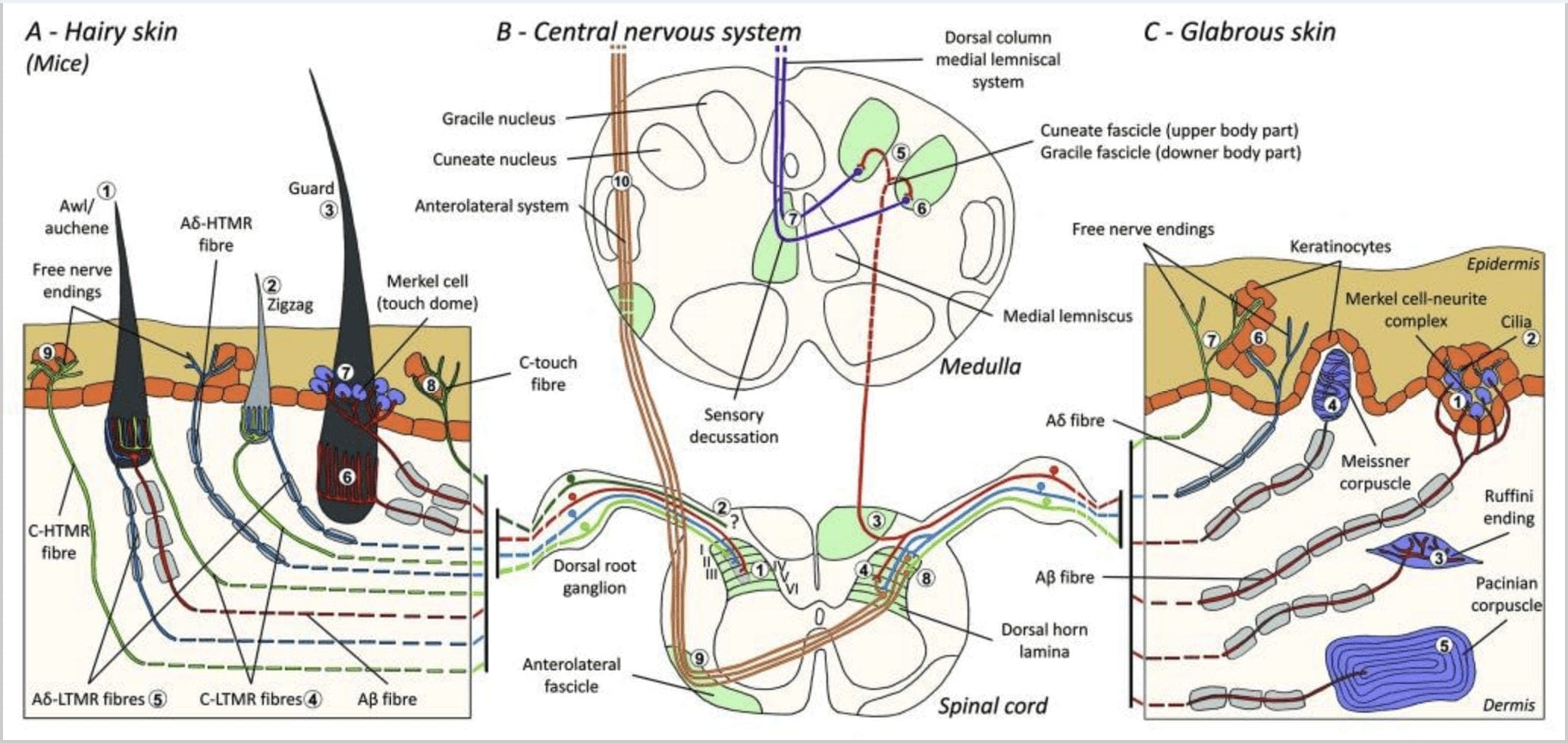
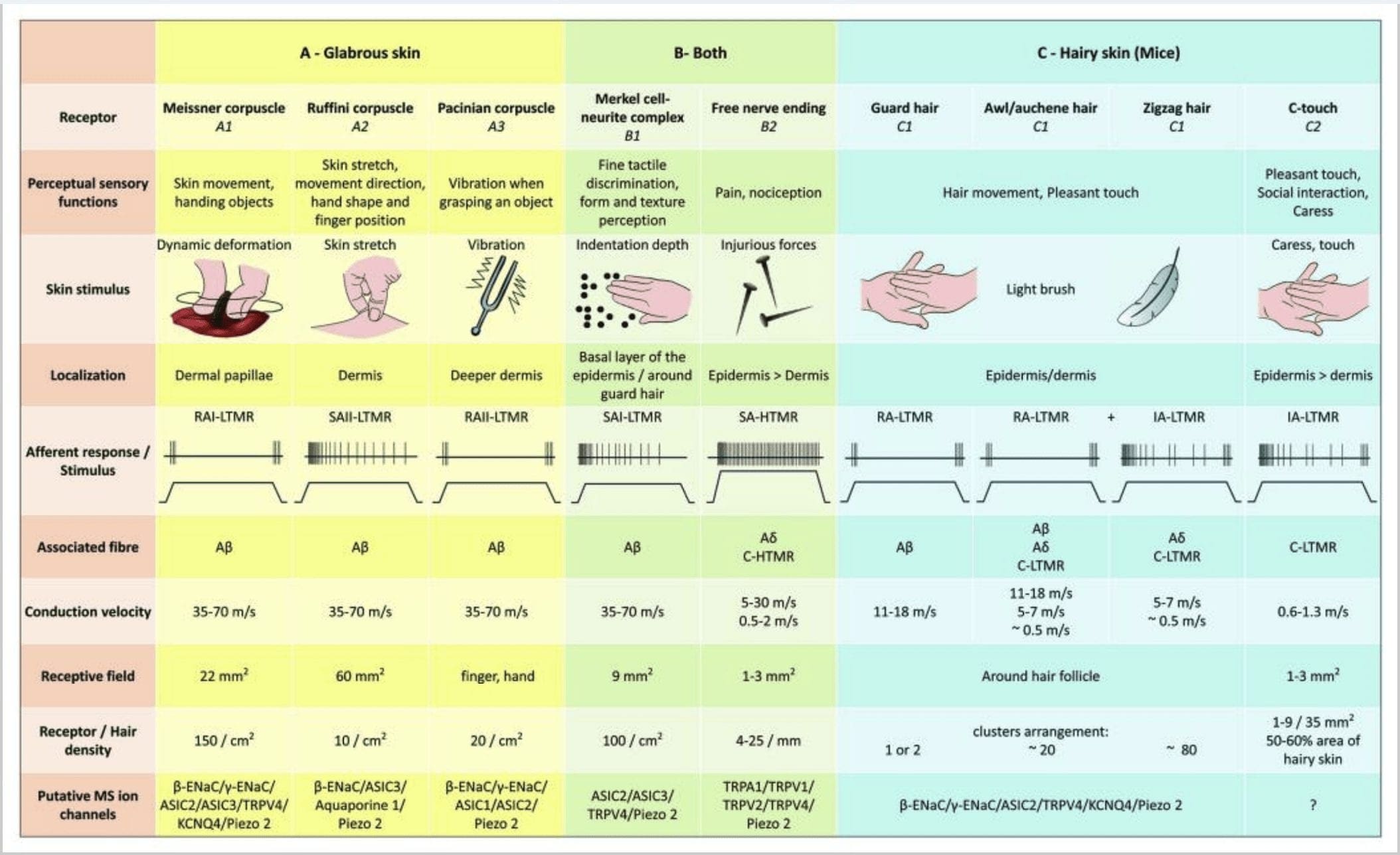

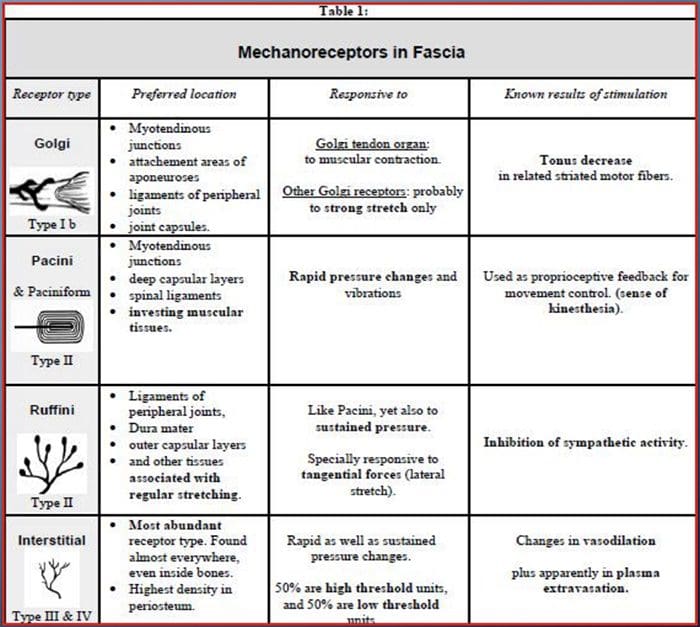 TEORIA BÓLU Z BRAMAMI
TEORIA BÓLU Z BRAMAMI
 DORSAL COLUMN MEDIAL LEMNISCAL PATHWAY
DORSAL COLUMN MEDIAL LEMNISCAL PATHWAY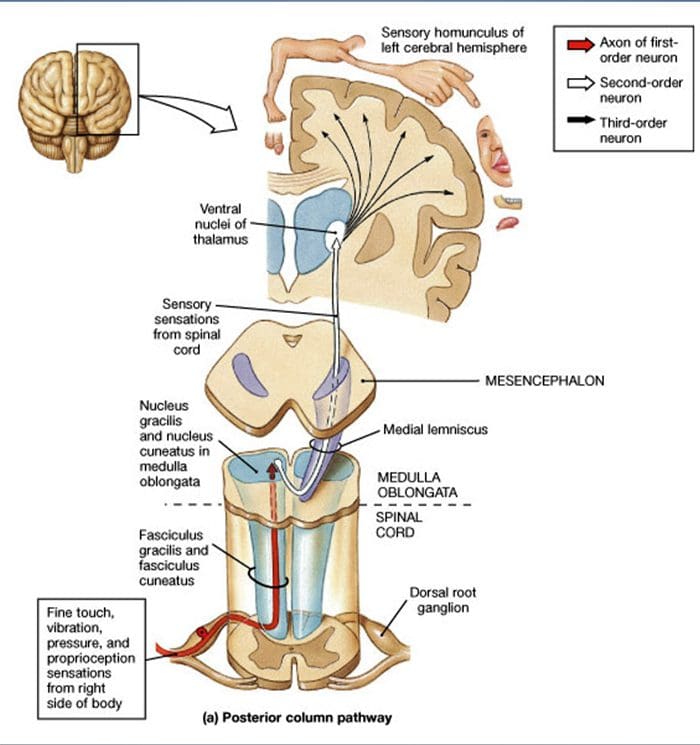 WYKORZYSTANIE PERIPHERAL MECHANORECEPTORÓW DO ZMIANY BÓLU POSTRZEGANIE
WYKORZYSTANIE PERIPHERAL MECHANORECEPTORÓW DO ZMIANY BÓLU POSTRZEGANIE
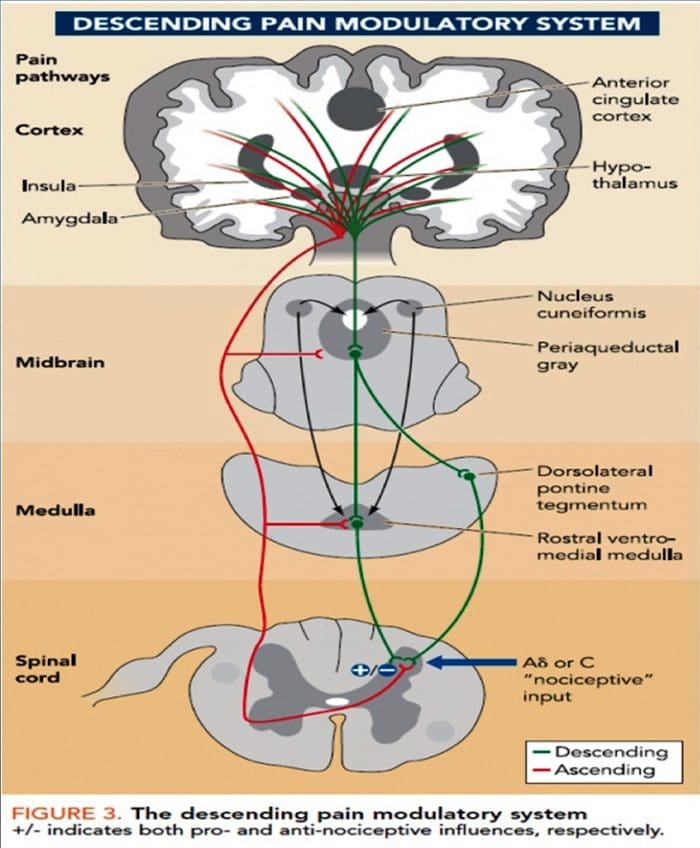
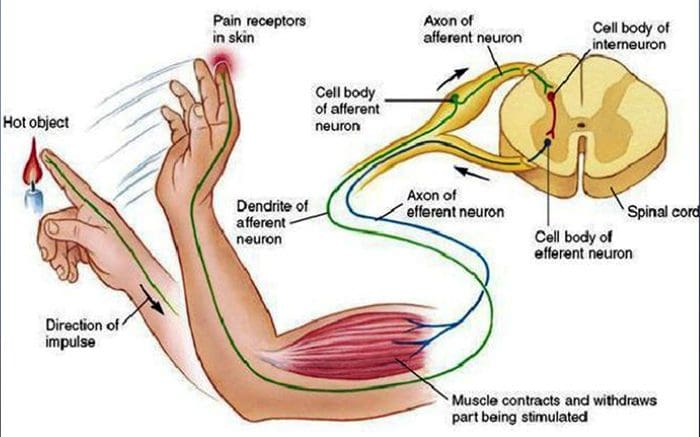
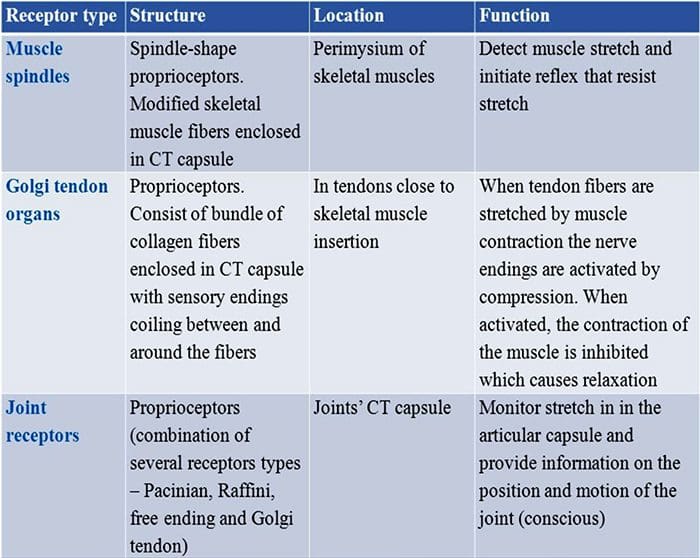
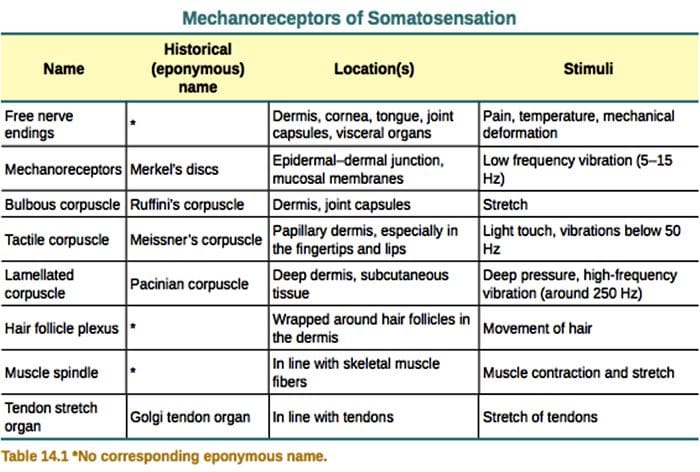 WIBRACJE
WIBRACJE
 LEKKI DOTYK
LEKKI DOTYK



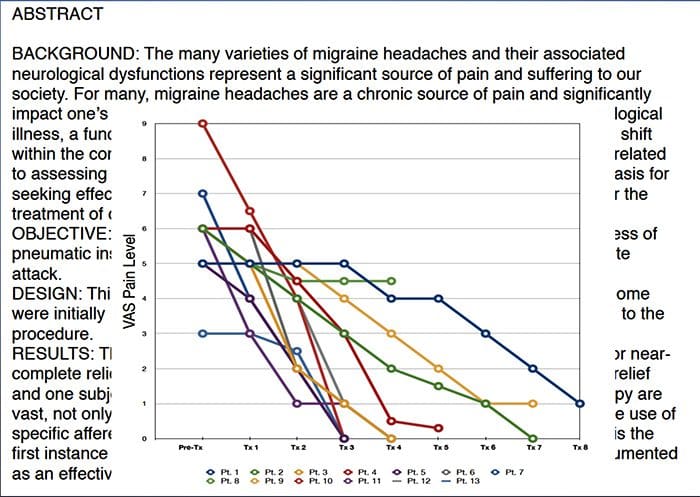

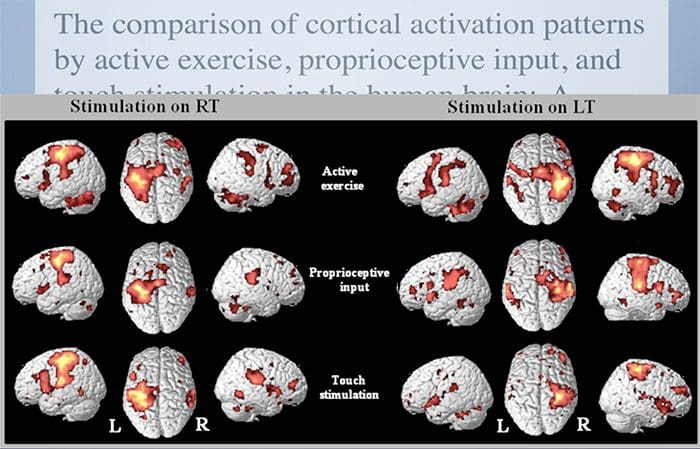
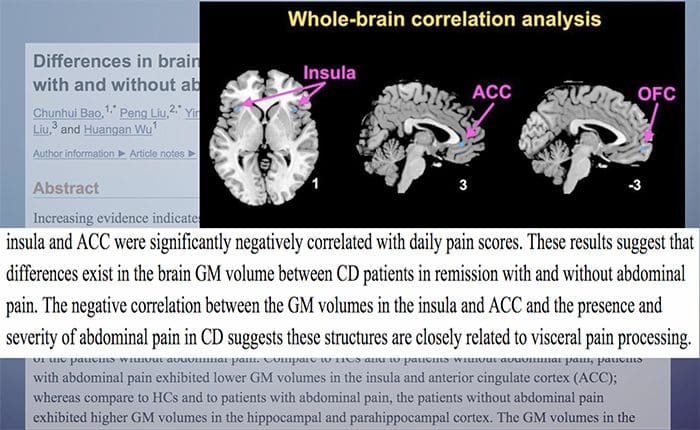

 CASE STUDY
CASE STUDY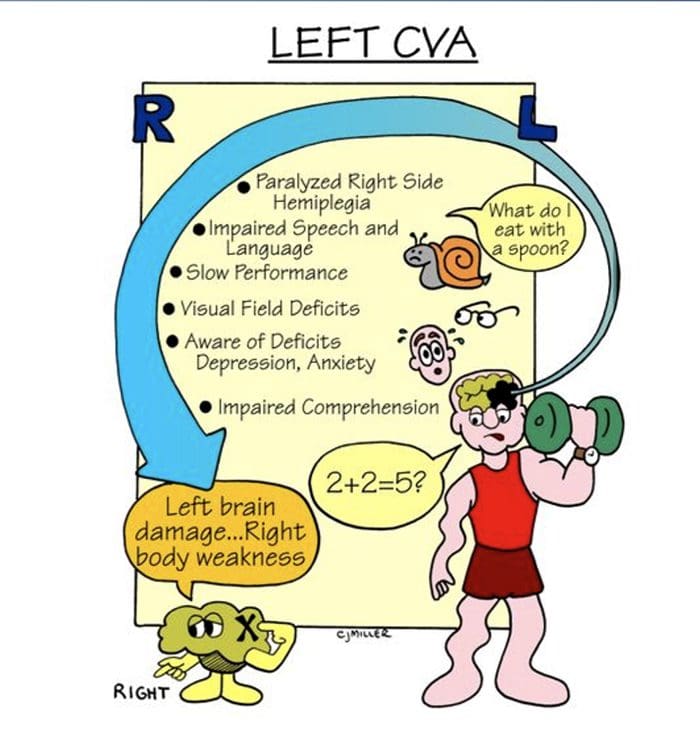 BADANIA NAUKOWE FIZYCZNE
BADANIA NAUKOWE FIZYCZNE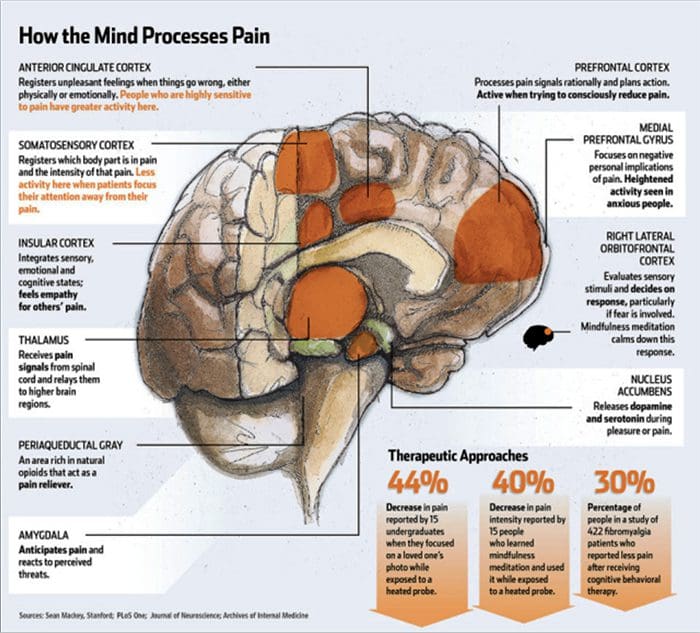 BADANIA NAUKOWE FIZYCZNE
BADANIA NAUKOWE FIZYCZNE
 PO DWÓCH DNIACH
PO DWÓCH DNIACH